Details of DPV and References
DPV NO: 280 July 1984
Family: Bromoviridae
Genus: Ilarvirus
Species: American plum line pattern virus | Acronym: APLPV
American plum line pattern virus
R. W. Fulton Department of Plant Pathology, University of Wisconsin, Madison, WI 53706, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
The disease was described by
Valleau (1932),
Cation (1941) and
Zeller & Milbrath (1942).
The virus was transmitted by sap and its properties described by
Kirkpatrick, Cheney & Lindner (1964) and
Paulsen & Fulton (1968).
Synonyms
- Plum line pattern virus (in part) (Rev. appl. Mycol. 44: 177)
- Peach line pattern virosis virus (Rev. appl. Mycol. 25: 218)
- Marmor pallidolimbatus, Prunus virus 10 (Rev. appl. Mycol. 21: 532)
- Peach line pattern virosis virus (Rev. appl. Mycol. 25: 218)
-
A virus with RNA-containing particles about 26, 28, 31 and 33 nm in diameter, sedimenting as four components. The virus is transmitted by inoculation of sap, more readily from experimental herbaceous hosts than from its natural woody hosts. The virus loses infectivity rapidly in sap unless an antioxidant is present. No vector known. Reported from USA and Canada.
Main Diseases
The virus causes ‘line pattern’ diseases (chlorotic lines and bands) in Prunus domestica and other plums, oriental flowering cherry (P. serrulata), peach (P. persica; Fig. 2) and other Prunus spp. (Kirkpatrick & Fulton, 1976). However, very similar symptoms are induced by apple mosaic virus and certain strains of prunus necrotic ringspot virus, neither of which is serologically related to American plum line pattern virus.
Geographical Distribution
Reported from the USA and Canada (Willison, 1945). It may, however, be more widely distributed because line pattern diseases have been reported from most countries where plums are grown, but the identity of the viruses present is not known for many of the earlier reports.
Host Range and Symptomatology
The virus has a wide host range; it infected 85 of 245 species inoculated with sap, including six woody rosaceous species (Paulsen & Fulton, 1968). It occurs naturally in many varieties of plum and other Prunus spp. (Kirkpatrick & Fulton, 1976).
-
Diagnostic species
- Cucumis sativus
(cucumber). May develop small (2 nm) chlorotic lesions in inoculated cotyledons, but the virus does not become systemic. - Nicotiana megalosiphon. Inoculated leaves develop chlorotic or diffusely
necrotic primary lesions. Systemic symptoms consist of a mild ringspotting or mottle
(Fig. 3).
- Physalis floridana. Inconspicuous chlorotic primary lesions followed by systemic chlorotic veinbanding, ringspotting or oak-leaf patterns.
- Prunus spp. Chlorotic lines, rings and oak-leaf patterns of varying severity (Fig. 2). In myrobalan and Shiro plum (P. cerasifera and P. americana x P. salicina) , some leaves may show ‘yellow net’ symptoms bordering the veins and veinlets. Ember plum reacts more severely than Shiro plum, which is commonly used as an indicator host (Fridlund, 1958).
- Vigna unguiculata ssp. cylindrica (catjang). Small reddish lesions within 3-4 days in primary leaves, provided that they are inoculated when about two-thirds expanded (Fig. 1). Systemic symptoms consist of a characteristic reddish etching of the veins of trifoliolate leaves, with stunting and curling (Fig. 4).
- Physalis floridana. Inconspicuous chlorotic primary lesions followed by systemic chlorotic veinbanding, ringspotting or oak-leaf patterns.
-
Propagation species
- Cultures may be maintained in Prunus spp. or in Vinca rosea. Young
leaves of V. rosea, or petals of Prunus, provide the most infective
inoculum. The virus may be propagated for purification in Nicotiana megalosiphon
or N. occidentalis.
Assay species
- Vigna unguiculata
spp. cylindrica has been used as a local lesion host. Many other leguminous species develop necrotic local lesions but in most species the lesions are few.
Strains
The isolate described by Kirkpatrick et al. (1964) infected a number of species (Gomphrena globosa, Lactuca sativa, Nicotiana glutinosa, N. rustica, Tithonia speciosa) that were apparently not susceptible to a Wisconsin isolate.
Transmission by Vectors
No vectors described.
Transmission through Seed
Seed transmission in Prunus has not been investigated. The virus was not transmitted through seed of Petunia hybrida, P. violacea or Phaseolus vulgaris (A. Q. Paulsen & R. W. Fulton, unpublished data).
Transmission by Dodder
Cuscuta campestris transmitted the virus from Nicotiana megalosiphon to Petunia hybrida, but C. subinclusa did not.
Serology
The virus is a moderately good immunogen in rabbits. Intramuscular injections twice a week of 1.8-2.0 mg of purified virus in Freund’s incomplete adjuvant resulted in antiserum titres of 1/1280-1/2560 after 6-7 weeks. Agar gel double diffusion tests showed a single line of precipitate with extracts of herbaceous hosts, but only ELISA was effective for detecting the virus in woody hosts (Fulton, 1982).
Relationships
The isolate of American plum line pattern virus described by Kirkpatrick et al. (1964) reacted strongly with antiserum to a Wisconsin isolate although their host ranges were not identical (Paulsen & Fulton, 1968). The properties and morphology of the virus suggest affinities with the ilarviruses (Fulton, 1983), but Paulsen & Fulton (1969) detected no serological relationship to three members of that group, prune dwarf, prunus necrotic ringspot and apple mosaic viruses, the last two of which also cause line pattern symptoms in plum. Barnett & Murant (1970) found no reaction between antiserum to American plum line pattern virus and raspberry bushy dwarf virus.
Stability in Sap
Undiluted sap of infected N. megalosiphon was not infective even when inoculated to catjang test plants immediately after preparation, but extracts made by grinding tissue in several volumes of phosphate buffer had some infectivity. Infectivity was stabilised for 4 h or more by grinding N. megalosiphon tissue in buffer containing 0.02 M 2-mercaptoethanol. In such extracts, assayed on catjang, the thermal inactivation point (10 min) of the virus was 66°C, and the dilution end-point was between 1/6400 and 1/12,800 (Paulsen & Fulton, 1968). The virus retained infectivity for several years when infected tissue of Petunia hybrida was dried over CaCl2 at 2°C.
Purification
(Paulsen & Fulton, 1969). Mature leaves of N. megalosiphon or N. occidentalis heavily inoculated 6-10 days previously with extracts from the same host, or from systemically infected leaves of catjang, are a suitable source of virus for purification. Homogenise the leaves in cold 0.02 M phosphate buffer, pH 8.0, containing 0.02 M 2-mercaptoethanol, at the rate of 1.5 ml/g tissue. Centrifuge the homogenate at low speed (1800 g for 0. 5 h) and adjust the supernatant liquid to pH 4.8 with 0.1 M citric acid. After holding at 4°C for 2 h, remove the green precipitate by centrifuging the extract at 1800 g for 20 min. Then adjust the supernatant liquid to pH 6.5-7.0 and centrifuge again at 1800 g for 0.5 h. Concentrate the virus from the clarified extract by centrifuging at 78,000 g for 3 h. Resuspend the pellets in 0.02 M EDTA, pH 6.2, and remove any insoluble material by low speed centrifugation. Repeat the high and low speed centrifugation cycle. The gamma globulin portion of antiserum to healthy N. megalosiphon has been used to remove remaining host protein (Paulsen & Fulton, 1969). This may also be done by centrifuging the preparation in sucrose gradients (7-25%) prepared in 0.03 M EDTA (Fulton, 1982). Yields of virus are about 2 mg/100 g fresh leaves. The particles disrupt quickly in water or a number of buffers, but retain morphological integrity and infectivity for several years in 0.03 M EDTA, pH 6.2, at 4°C. Purified virus retained infectivity longer in ammonium phosphate (pH 8.0) than in sodium or potassium phosphate.
Properties of Particles
Preparations of virus particles sediment as four components (T, M, B1 and B2) in sucrose density gradients (Fig. 5). The separated components were only weakly infective, mixtures of all four components were the most infective; however the components were probably not completely separated.
Sedimentation coefficients (s20, w): about 95, 100, 114 and 125 S in comparison with known viruses as standards (Fulton, 1982).
A260/A280: 1.67 (unfractionated virus).
Particle Structure
The nucleoprotein particles are quasi-isometric (Fig. 6) and those from the four bands in sucrose density gradients have diameters of 26, 28, 31 and 33 nm, from the uppermost to the lowest band.
Particle Composition
Nucleic acid: No information. However, by analogy with similar multicomponent viruses the nucleic acid is probably single-stranded RNA. If so the A260/A280 ratio would suggest that the RNA comprises about 16% of the particle weight.
Protein: No information.
Relations with Cells and Tissues
No information.
Notes
American plum line pattern virus is not serologically related to either prunus necrotic ringspot or apple mosaic viruses and does not cause mosaic in apple. All three of these (or strains of them) may cause line pattern in plum. The specific designation ‘American plum line pattern virus’ was suggested by Seneviratne & Posnette (1979). Other terms e.g. apricot line pattern (Schuch, 1961), are presumed to refer to apple mosaic or prunus necrotic ringspot viruses.
Figures

Primary lesions in Vigna unguiculata ssp. cylindrica (catjang) 4 days after inoculation.

Systemic symptoms in peach.

Systemically infected leaf of Nicotiana megalosiphon.

Systemic symptoms in first trifoliolate leaf of V. unguiculata spp. cylindrica.

Distribution of sedimenting components T, M, B1 and B2 after centrifugation in a density gradient of sucrose (7-25%) for 4 h at 25,000 rev/min in a Beckman SW 25.1 rotor.
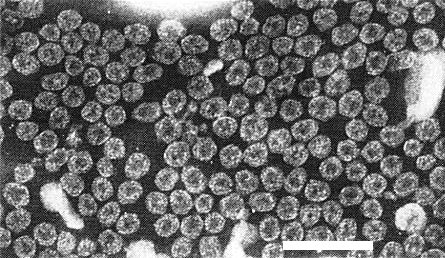
Purified virus particles, fixed with glutaraldehyde and negatively stained with sodium phosphotungstate. Bar represents 100 nm.
References list for DPV: American plum line pattern virus (280)
- Barnett & Murant, Ann. appl. Biol. 65: 435, 1970.
- Cation, Phytopathology 31: 1004, 1941.
- Fridlund, Pl. Dis. Reptr 42: 1051, 1958.
- Fulton, Phytopathology 72: 1345, 1982.
- Fulton, CMI/AAB Descr. Pl. Viruses, 275, 3 pp., 1983.
- Kirkpatrick, Cheney & Lindner, Pl. Dis. Reptr 48: 616, 1964.
- Kirkpatrick & Fulton, in Virus Diseases and Noninfectious Disorders of Stone Fruits in North America. U.S. Dept. Agric., Agric. Handbook 437, 166, 1976.
- Paulsen & Fulton, Phytopathology 58: 766, 1968.
- Paulsen & Fulton, Ann. appl. Biol. 63: 233, 1969.
- Schuch, Z. PflKrankh. 68: 10, 1961.
- Seneviratne & Posnette, Ann. appl. Biol. 65: 115, 1970.
- Valleau, Res. Bull. Ky agric. Exp. Stn 327: 89, 1932.
- Willison, Phytopathology 35: 991, 1945.
- Zeller & Milbrath, Phytopathology 32: 634, 1942.