Details of DPV and References
DPV NO: 281 July 1984
Family: Bromoviridae
Genus: Ilarvirus
Species: Spinach latent virus | Acronym: SpLV
Spinach latent virus
L. Bos Research Institute for Plant Protection, Binnenhaven 12, P. O. Box 9060, 6700 GW Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Bos, Huttinga & Maat (1980),
Van der Meer (1968), and
Stefanac (1978).
Synonym
- GE36 virus (Rev. Pl. Path. 49: 2522b)
-
A virus with quasi-isometric particles c. 27 nm in diameter, which sediment as three components. Found only in spinach from which it is readily transmissible by inoculation of sap to a number of herbaceous hosts, in most of which it is symptomless. Seed-borne with high frequency in a number of hosts. Vector unknown. Reported from Yugoslavia and the Netherlands.
Main Diseases
Occurs as a seed-borne infection of spinach, in which it induces no symptoms (17 cultivars tested by Bos et al., 1980) or only transient symptoms (Stefanac, 1978). Reported infection of apple and pear (GE36 virus; Van der Meer, 1968) may be erroneous (Van der Meer, 1974), the virus having probably been a contaminant of the seed of test plants (GE36 virus could not be returned to apple and was seed transmitted in Chenopodium quinoa). The virus is of potential importance because of its probable prevalence, symptomless infection and high frequency of seed transmission in many plant species.
Geographical Distribution
Reported only from Yugoslavia (Stefanac, 1978; Stefenac & Wrischer, 1983) and the Netherlands (Bos et al., 1980), but also detected in seed of spinach from other countries.
Host Range and Symptomatology
The virus infected 25 out of 48 dicotyledonous plant species in 10 out of 12 families, most of them systemically, but normally without producing symptoms (Bos et al., 1980; Stefenac & Wrischer, 1983). Plants are more readily infected and symptoms more obvious at 20°C than at 23°C (Stefenac & Wrischer, 1983).
-
Diagnostic species
- Beta vulgaris
(beet). Most cultivars tested developed small chlorotic or etched rings on inoculated leaves (Fig. 4), normally transient. Symptomless systemic infection. - Celosia cristata. Symptomless local and systemic infection.
- Chenopodium amaranticolor. Numerous pin-point desiccated (on lower leaves) or chlorotic (on upper leaves) local lesions c. 3 days after inoculation (Fig. 1); no systemic infection.
- Chenopodium quinoa. A moderate number of chlorotic local lesions appear 3 to 4 days after inoculation; they are often vague and transient and in leaves inoculated young are accompanied by yellowish mottling (Fig. 2). Systemic symptoms appear 7 to 10 days after inoculation with slight leaf rolling followed by chlorotic vein banding, yellow mottling, growth reduction, and sometimes desiccation of young leaves; symptom severity depends on inoculum dose (Fig. 3).
- Cucumis sativus (cucumber) ‘Gele Tros’. Chlorotic local lesions, usually few and often transient. With the GE36 isolate, infection becomes systemic but is symptomless. This species is useful for distinguishing the virus from cucumber mosaic virus and tobacco rattle virus which may also be seed-borne in spinach.
- Nicotiana clevelandii. N. glutinosa, N. rustica, N. tabacum ‘Samsun’, ‘White Burley’ and ‘Xanthi’. All develop symptomless systemic infection. At low temperatures N. glutinosa and N. tabacum may show systemic mild irregular fine vein chlorosis or necrosis on some leaves (Stefenac & Wrischer, 1983).
- Phaseolus vulgaris (French bean) ‘Bataaf’. Distinct necrotic local lesions c. 1 mm in diameter appear 3 to 4 days after inoculation (Fig. 5). No systemic infection.
- Chenopodium amaranticolor. Numerous pin-point desiccated (on lower leaves) or chlorotic (on upper leaves) local lesions c. 3 days after inoculation (Fig. 1); no systemic infection.
-
Propagation species
- Chenopodium quinoa
and Nicotiana spp.Assay species
- Chenopodium amaranticolor, C. quinoa
and Phaseolus vulgaris.
Strains
No obvious differences were observed between isolates from different lots of spinach seed. Spinach isolates were not found to differ appreciably from GE36 isolate (Van der Meer, 1968) in host range and symptoms, serology or biophysical characteristics, though GE36 tended to be less virulent and less easily seed-transmitted. The Yugoslavian isolate (Stefenac, 1978) was reported prior to the Dutch one and, though uncharacterised, was coincidentally also named spinach latent virus. It was later found to be identical with the Dutch virus in biological properties and gave reactions of identity in agar gel diffusion tests with antiserum to the Dutch isolate. The more severe symptoms observed in Yugoslavia may have been caused by lower temperatures (Stefenac & Wrischer, 1983).
Transmission by Vectors
No vectors are known. Not transmitted by Myzus persicae (Bos et al., 1980).
Transmission through Seed
Frequencies of seed transmission are high. Bos et al. (1980) found more than 50% infection in some commercial seed of spinach, over 50% in seed from infected plants of Celosia cristata, over 90% in Chenopodium quinoa and Nicotiana tabacum and c. 30% in N. rustica. Stefenac & Wrischer (1983) reported 90% seed transmission in N. clevelandii and 95% in N. megalosiphon. With the GE36 isolate, transmission frequencies were lower (Van der Meer, 1974; Bos et al., 1980). Transmission by pollen to seed of C. quinoa and spinach, and to pollinated plants of spinach, is also reported (Stefenac, 1978; Stefenac & Wrischer, 1983). The virus is present in the embryo (Bos et al., 1980).
Serology
Weakly immunogenic in rabbits. Antisera with titres of 1/64 have been prepared to the GE36 isolate (Maat & Vink, 1971) and to the type isolate from spinach (Bos et al., 1980). The antisera did not react with virus-containing crude sap. With purified virus two precipitin lines are produced in agar in 1% NaCl but only one line in agar in 0.15 M phosphate-citric acid buffer, pH 7 (Bos et al., 1980; Stefenac & Wrischer, 1983).
Relationships
The physico-chemical properties of the virus place it in the ilarvirus group, though the particles did not react with antisera to apple mosaic, elm mottle, lilac ring mottle, prunus necrotic ringspot or tobacco streak viruses. However, ilarviruses in general are poorly immunogenic, and serological relationships are limited to members of the same subgroup. The fifth RNA species is unusual among ilarviruses and other viruses with tripartite genomes but it has not been investigated separately and may be an artifact of purification.
Stability in Sap
In sap of Chenopodium quinoa the thermal inactivation point (10 min) is c. 60-65°C, longevity at room temperature 6-8 days (> 13 days in one test) and dilution end-point between 10-3 and 10-4 (Bos et al., 1980; Stefenac & Wrischer, 1983). The virus survived at least 5-6 years after drying and storage of infected leaves over CaCl2 at 4°C (Stefenac & Wrischer, 1983) and in seeds of Chenopodium quinoa (L. Bos, unpublished results), and at least 5 years in seeds of spinach kept at room temperature (Stefenac & Wrischer, 1983).
Purification
The following method is effective (Bos et al., 1980). Homogenise infected leaves of Chenopodium quinoa, after storing them at 4°C overnight, in 100 g portions in a blender in 300 ml 0.018 M phosphate-citric acid buffer, pH 7, containing 0.1% thioglycollic acid. Filter through cheesecloth. Bring filtrate to 8% with n-butanol, stir for 45 min and leave for 30 min at 4°C. Centrifuge the homogenate for 10 min at low speed (16,000 g). Concentrate the virus from the upper phase by two cycles of differential centrifugation. Purify further by two cycles of sucrose density-gradient centrifugation, or by isopycnic centrifugation in Cs2SO4.
Properties of Particles
Purified preparations contain three centrifugal components (Bos et al., 1980). Sedimentation coefficients, s°20,w, in 0.018 M phosphate-citric acid buffer, pH 7: 87, 98 and 103 S.
Buoyant density of all components: 1.269 g cm-3 in Cs2SO4.
Particle Structure
Particles are labile and difficult to locate in crude sap without fixation before staining with phosphotungstate for electron microscopy. In purified preparations they are irregularly shaped, c. 27 nm in diameter and some are bacilliform (Fig. 6).
Particle Composition
Nucleic acid: RNA, single-stranded. Unfractionated preparations contain five RNA species with M. Wt of 1.30 x 106 (RNA-1), 1.18 x 106 (RNA-2), 0.91 x 106 (RNA-3), 0.35 x 106 (RNA-4) and 0.27 x 106 (RNA-5), estimated by polyacrylamide gel electrophoresis under partially denaturing conditions with cowpea chlorotic mottle virus RNA species as internal markers. For infection the three larger RNA species are required plus one or both of the two small components or the coat protein (Bos et al., 1980).
Protein: The virus particles contain one type of polypeptide of M. Wt 28,000 (Bos et al., 1980).
Relations with Cells and Tissues
In infected plants of C. quinoa and B. vulgaris no inclusion bodies were observed in epidermal cells by light microscopy, but electron microscopy of ultrathin sections revealed groups of vesicles surrounded by a unit membrane in the cytoplasm and small vacuoles of leaf parenchyma cells (Stefenac & Wrischer, 1983).
Notes
During investigations on the virus, cucumber mosaic virus was detected four times and tobacco rattle virus once in seed samples of spinach. These two viruses can easily be distinguished from spinach latent virus. Cucumber mosaic virus and tobacco rattle virus are rarely systemic in Chenopodium quinoa. Cucumber mosaic virus causes characteristic local lesions in C. quinoa and infects cucumber and Nicotiana species systemically. Tobacco rattle virus causes obvious local lesions in cucumber cotyledons and some Nicotiana species and gives erratic systemic infections in N. tabacum (Bos et al., 1980).
Figures

Chenopodium amaranticolor with numerous local lesions 7 days after inoculation.

Chenopodium quinoa with some desiccated local lesions and chlorotic mottling 14 days after inoculation.

Chenopodium quinoa with severe systemic symptoms 28 days after inoculation: (left to right) high dose of virus, low dose of virus, and uninoculated control.

Beta vulgaris (beet) with local chlorotic rings 13 days after inoculation.
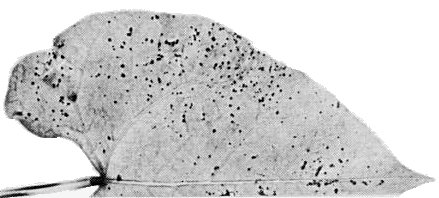
Phaseolus vulgaris (French bean) ‘Bataaf’ with necrotic local lesions 20 days after inoculation.

Particles of purified virus stained in 2% phosphotungstate, pH 6.5, in water. Bar represents 100 nm. (Photo courtesy Dr H. Huttinga.)
References list for DPV: Spinach latent virus (281)
- Bos, Huttinga & Maat, Neth. J. Pl. Path. 86: 79, 1980.
- Maat & Vink, Neth. J. Pl. Path. 77: 73, 1971.
- Stefenac, Acta bot. croat. 37: 39, 1978.
- Stefanac & Wrischer, Acta bot. croat. 42: 1, 1983.
- Van der Meer, TagBer. dt. Akad. LandwWiss. Berl. 97:27, 1968.
- Van der Meer, Jversl. Inst. plziektenk. Onderz., 1973: 52, 1974.