Details of DPV and References
DPV NO: 283 July 1984
Family: Tombusviridae
Genus: Aureusvirus
Species: Maize white line mosaic virus | Acronym: MWLMV
Maize white line mosaic virus
G. A. de Zoeten Department of Plant Pathology, University of Wisconsin, Madison, WI 53706, USA
B. B. Reddick Department of Entomology and Plant Pathology, University of Tennessee, Knoxville, TN 37901, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Boothroyd & Israel (1980).
A virus with 35 nm isometric particles sedimenting as a single component, and containing a single ssRNA species. The virus is transmitted in a soilborne manner, probably by fungi, but has not been transmitted by mechanical inoculation. It causes severe stunting and a bright mosaic in maize (corn) but has a very narrow host range, infecting only a few species of monocotyledonous plants. Reported from USA, France and Italy. A satellite-like virus with isometric particles c. 17 nm in diameter is associated with some isolates.
Main Diseases
Causes severe mosaic and stunting of dent and sweet corn types of maize (Zea mays) (Boothroyd & Israel, 1980). Cobs do not fill out, are mis-shapen and contain few kernels. Yield losses range from 5 to 45% in infected fields. The disease seems to be associated with wet, poorly drained soils.
Geographical Distribution
The disease has been reported from East and North Central USA. A similar disease occurs in France and Italy (Gingery & Louie, 1984; H. Lapierre and D. T. Gordon, personal communications).
Host Range and Symptomatology
The virus has not been transmitted mechanically and has been found naturally in only a few members of the Gramineae. In addition to maize it infects Digitaria sanguinalis, Panicum dichotomiflorum. Setaria faberi, S. viridis and Triticum aestivum (de Zoeten et al., 1980; Grau et al., 1981; Louie et al., 1982).
-
Diagnostic species
- Zea mays
(maize, dent corn, sweet corn). Infected plants are severely stunted, the leaves showing a bright mosaic consisting of white, yellow and yellow-green rectangular to linear (1-2 cm) patches and stripes (Fig. 1, Fig. 2). The symptoms are not confined to whorl leaves (as are those of maize dwarf mosaic virus), but are present in all leaves and persist after pollination. However, the virus can be present in maize and other hosts without inducing symptoms (Louie et al., 1982).Propagation and assay species
- Zea mays
.
Strains
All isolates studied so far seem serologically identical.
Transmission by Vectors
The virus is transmitted in a soilborne manner. The vector is unknown but could be a fungus because transmission was not abolished by air drying infective soil and was prevented or decreased by treating the soil with benomyl but not with carbofuran, an insecticide/nematicide (Louie et al., 1982). Moreover, structures similar to the resting sporangia of Polymyxa and Olpidium were found in roots of infected plants (Grau et al., 1981; Louie, Gordon & Lipps, 1981). A satellite-like virus associated with some isolates is also soil-transmitted (Gingery & Louie, 1984).
Transmission through Seed
Virus antigen has been detected in pollen, silks and kernels of infected maize plants (Louie et al., 1982). Although the possibility of seed transmission in maize cannot be excluded it must be less than 0.01% (Louie et al., 1982).
Serology
The virus is strongly immunogenic in rabbits; antisera with titres of 1/2048 are readily obtainable (de Zoeten, 1980). Microprecipitin and gel double diffusion tests, and ELISA, are all useful for detecting antigen in extracts from infected plants.
Relationships
Maize white line mosaic virus has no obvious similarities to any other plant virus. Serological tests failed to detect any relationship of the virus to maize chlorotic dwarf, maize chlorotic mottle or maize rayado fino viruses (Louie et al., 1982). The satellite-like virus found in some isolates is serologically unrelated to maize white line mosaic virus or to the satellites associated with tobacco necrosis and panicum mosaic viruses (Gingery & Louie, 1984).
Stability in Sap
Not known. Leaf dips (Fig. 3) from infected tissue show particles similar to those seen in purified preparations, indicating good stability.
Purification
Two methods are reported:
(1) Louie et al. (1982). Homogenise infected maize leaves in 3 vol. phosphate-citrate buffer (0.2 M K2HPO4 adjusted to pH 6.0 with 0.1 M citric acid) containing 0.5% 2-mercaptoethanol. Emulsify leaf homogenates with 0.33 vol. chloroform and centrifuge at 10,500 g for 15 min. Collect the aqueous phase, centrifuge at 125,000 g for 90 min at 10°C and resuspend the pellets in phosphate-citrate buffer. Purify further by centrifugation in 10-40% linear sucrose gradients in the same buffer (2-5 h at 200,000 g). Collect the virus band, dilute threefold, add CsCl to a final concentration of 1.33 g/ml, and centrifuge at 125,000 g for 18-24 h. Collect the virus zone and concentrate the virus by high speed centrifugation. Resuspend pellets in phosphate-citrate buffer. Yields are 400-600 mg virus/kg tissue.
(2) de Zoeten et al. (1980). Freeze virus-infected tissues at -20°C. Triturate each 1 kg tissue in a Waring blender with 1 litre extraction buffer (0.02 M phosphate buffer, pH 7.2, containing 0.02 M diethyldithiocarbamate and 0.02 M mercaptoethanol). Press the triturate through four layers of cheesecloth, mix with 0.25 vol. chloroform/butanol (1:1) and centrifuge for 10 min at 10,000 g. Collect the upper aqueous phase and centrifuge for 90 min at 78,000 g. Resuspend the pellet in 0.02 M phosphate buffer (pH 7.2) and centrifuge the suspension for 5 min at 8000 g. Layer the supernatant fluid over 20% sucrose (5 ml in Spinco type 40 rotor tubes) and centrifuge for 2 h at 105,500 g. Purify further by centrifugation in 10-40% (w/v) sucrose density gradients (3 h at 82,000 g). Prepare all sucrose solutions in 0.02 M phosphate buffer pH 7.2. Yields are 200-400 mg virus/kg tissue.
Properties of Particles
In cultures free from the satellite-like virus, the particles sediment as a single component in both sucrose density gradient (Fig. 4) and CsCl equilibrium centrifugation. Particles of the satellite-like virus form an additional sedimenting component.
Sedimentation coefficient (s20,w): 117 S. Particles of the satellite-like virus sediment at 47 S (Gingery & Louie, 1984).
Buoyant density in CsCl: 1.353 g/cm3 measured in an analytical centrifuge with tobacco mosaic virus as an internal standard (de Zoeten et al., 1980). Particles of the satellite-like virus have a buoyant densiy of 1.355 g/cm3 (Gingery & Louie, 1984).
A260(0.1%; 1 cm): 3.9; A260/A280: 1.7; both values corrected for light-scattering (de Zoeten et al., 1980).
Particle Structure
Virus particles are isometric with a diameter of 35 nm (Fig. 3, Fig. 5). Particles in sectioned tissue prepared for electron microscopy measured 24-28 nm (Fig. 6). The particles of the satellite-like virus are c. 17 nm in diameter (Gingery & Louie, 1984).
Particle Composition
Nucleic acid: RNA, single-stranded, Mr approximately 1.25 x 106, determined by polyacrylamide gel electrophoresis under denaturing conditions with brome mosaic virus RNA species as standards. The RNA comprises 21% of the particle weight. Molar percentages of nucleotides: G = 30.0; A = 19.7; C = 26.0; U = 24.3. The RNA of the satellite-like virus is single-stranded and has a M. Wt of 0.4 x 106 (R.E. Gingery, personal communication).
Protein: Particles contain a single protein structural unit with a Mr variously reported to be 32,000 (de Zoeten et al., 1980) or 35,300 (Louie et al., 1982). The particles of the satellite-like virus contain a protein of Mr 24,700 (Gingery & Louie, 1984).
Relations with Cells and Tissues
Electron microscopy of thin sections of infected sweet corn leaves revealed large concentrations of virus particles in virtually all tissues including xylem elements (de Zoeten et al., 1980). The size of the virus particles in tissues ranged from 24 to 28 nm with a predominant size of ~25 nm. The virus particles were easily recognisable because of their dark-staining characteristics whether present singly or in large aggregates (Fig. 6). Although some regularity in virus particle arrangement could be seen occasionally, paracrystalline virus structures were not observed. Virus particles were seen only in the cytoplasm. Vesiculation of the cytoplasm and increased amounts of endoplasmic reticulum, suggestive of a replication complex, often were observed in areas containing virus particles. Large polyribosomes and/or a spiral arrangement of virus particles seemed to be a common feature of these areas. Three types of virus image were seen in tissue sections: dark, heavily stained particles; lightly stained particles; and (occasionally) particles that seemed to have little or no stain in the centre.
Notes
The failure to transmit this virus mechanically has hampered further research, specifically in relation to its possible fungal vector. The virus is so different in size (35 nm) from most other small isometric viruses infecting maize, and is in such exceptionally high concentrations in the tissue, that it can be diagnosed by electron microscopy of leaf extracts. No other virus known in maize gives symptoms like those of maize white line mosaic virus. Although the early symptoms of maize dwarf mosaic virus infection are similar, they remain confined to the whorl leaves.
Figures

Systemic symptoms in leaves of maize.

A close-up of white line and mosaic symptoms in a maize leaf.
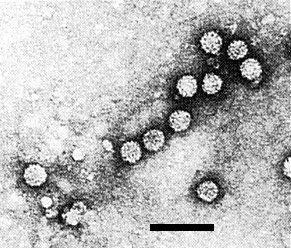
Virus particles from a leaf dip. Bar represents 100 nm.
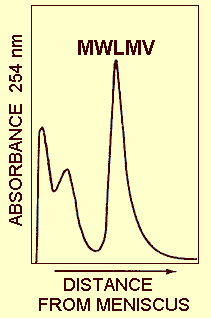
A sucrose density gradient profile obtained by centrifuging crude virus preparations through a 10-40% sucrose gradient. Note the single virus peak.
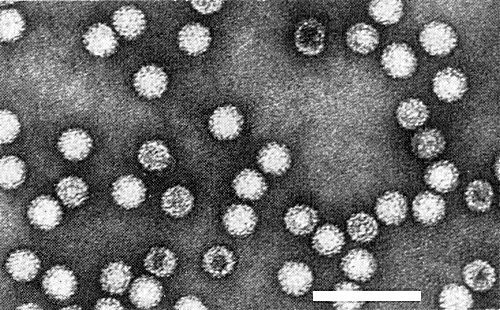
Virus particles recovered from sucrose gradients. Bar represents 100 nm.

Virus particles (V) in a cytoplasmic inclusion from a sweet corn leaf mesophyll cell. Note areas with vesicles (Ve) (short arrowheads) and the spiral arrangement of polyribosomes (long arrows). Ch = chloroplast; M = mitochondrion. Bar represents 500 nm.
References list for DPV: Maize white line mosaic virus (283)
- Boothroyd & Israel, Pl. Dis. 64: 218, 1980.
- de Zoeten, Arny, Grau, Saad & Gaard, Phytopathology 70: 1019, 1980.
- Gingery & Louie, Phytopathology 74: 808, 1984.
- Grau, de Zoeten, Arny, Saad & Gaard, Phytopathology 71: 220, 1981.
- Louie, Gordon & Lipps, Phytopathology 71: 1116, 1981.
- Louie, Gordon, Knoke, Gingery, Bradfute & Lipps, Pl. Dis. 66: 167, 1982.