Details of DPV and References
DPV NO: 289 July 1984
Family: Betaflexiviridae
Genus: Carlavirus
Species: Honeysuckle latent virus | Acronym: HnLV
Honeysuckle latent virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
F. A. Van der Meer Research Institute for Plant Protection, Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Van der Meer, Maat & Vink (1980) and Brunt, Phillips &
Thomas (1980).
- Synonym
- Lonicera latent virus (Rev. Pl. Path. 56, 959; Rev. Pl. Path.
60, 345)
- A virus with a monopartite RNA genome and slightly flexuous filamentous particles c. 650 nm long, commonly infecting honeysuckles (Lonicera japonica and L. periclymenum) in Europe. Transmitted in the non-persistent manner by aphids and by mechanical inoculation of sap.
Main Diseases
Infection is common, but usually symptomless, in many cultivars of Lonicera japonica and L. periclymenum (Caprifoliaceae). The virus, however, has not been found infecting L. nitida or the following commonly-grown members of the same family: Leycesteria formosa, Sambucus nigra, Symphoricarpos albus, Viburnum lantana and Weigela sp. Infected L. japonica and L. periclymenum plants sometimes also contain cucumber mosaic or tobacco leaf curl viruses (Van der Meer et al., 1980; Brunt et al., 1980).
Geographical Distribution
Reported only from Britain (Brunt et al., 1980) and the Netherlands (Van der Meer et al., 1980).
Host Range and Symptomatology
The virus can be transmitted easily by mechanical inoculation of sap to its two natural hosts and to nine other species in the Amaranthaceae, Chenopodiaceae, Cucurbitaceae and Solanaceae. The virus failed to infect any of seven species in the Aizoaceae, Cruciferae, Papilionaceae and Salicaceae (Brunt et al., 1980; Van der Meer et al., 1980).
- Diagnostic species
- Chenopodium amaranticolor
and C. murale. Chlorotic lesions in inoculated leaves (Fig. 1); no systemic infection. - C. quinoa. Chlorotic lesions in inoculated leaves (Fig. 1); some
isolates also induce chlorotic spotting of systemically infected leaves (Fig. 2).
- Nicotiana clevelandii and N. glutinosa. Faint circular chlorotic lesions in inoculated leaves (Fig. 3); diffuse chlorosis or chlorotic vein-banding in systemically infected leaves.
- N. megalosiphon. Circular chlorotic lesions in inoculated leaves (Fig. 4); chlorotic banding and necrosis of veins in systemically infected leaves (Fig. 5).
- Nicotiana clevelandii and N. glutinosa. Faint circular chlorotic lesions in inoculated leaves (Fig. 3); diffuse chlorosis or chlorotic vein-banding in systemically infected leaves.
- Propagation species
- Nicotiana clevelandii
and N. megalosiphon are useful for maintaining cultures and as sources of virus for purification.- Assay species
- Nicotiana megalosiphon, Chenopodium murale
and C. quinoa are moderately good local lesion assay hosts.
Strains
None reported.
Transmission by Vectors
Transmitted in the non-persistent manner by the aphid Hydaphis foeniculi, but not by Myzus persicae (Brunt et al., 1980; Van der Meer et al., 1980).
Transmission through Seed
No seed transmission detected in Lonicera periclymenum (Van der Meer et al., 1980).
Serology
The virus is a good immunogen; antiserum with a titre of up to 1/16,384 in tube precipitin tests was obtained 30 days after injecting a rabbit intramuscularly with c. 3 mg of virus emulsified with Freund’s complete adjuvant (Brunt et al., 1980). Immunosorbent electron microscopy and enzyme linked immunosorbent assay can be used for rapid virus detection and identification (A. A. Brunt & F. A. Van der Meer, unpublished information).
Relationships
Isolates from the Netherlands and Britain are serologically indistinguishable (Brunt et al., 1980). The virus is serologically distantly related to poplar mosaic virus (serological differentiation indices of 6-8) and shallot latent virus (serological differentiation index of 4), but is apparently unrelated to 13 other carlaviruses (Bos et al., 1978; Van der Meer et al., 1980; Brunt et al., 1980). Although related to poplar mosaic virus, honeysuckle latent virus does not infect Populus euramericana (Van der Meer et al., 1980).
Stability in Sap
Most isolates are stable in Nicotiana clevelandii or N. megalosiphon leaf extracts. The thermal inactivation point (10 min) is between 75 and 85°C, the dilution end-point between 10-3 and 10-4 and the longevity in vitro between 2 and 8 days at 20°C and from 32 to 64 days at 2°C (Van der Meer et al., 1980; Brunt et al., 1980).
Purification
The virus can be readily purified from Nicotiana clevelandii or N. benthamiana. The two following procedures have been used:
1. Extract leaves (1 g/3 ml) in 0.067 M phosphate buffer at pH 7.2 containing 0.1 M thioglycollate, add n-butanol slowly to 8% (v/v) and, after shaking for 30 min, subject the extract to one or two cycles of differential centrifugation (20 min at 12,000 g; 75 min at 65,000 g). Resuspend sedimented virus in neutral 0.033 M phosphate buffer (1 ml/20 g leaf tissue), and purify further by chromatography on columns of 700 Å (120-200 mesh) controlled-pore glass beads. This procedure yields 45-80 mg virus/kg leaf tissue (Brunt et al., 1980).
2. Homogenise leaves (1 g/5 ml) in cold 0.1 M Tris buffer at pH 9 containing 0.02 M sodium diethyldithiocarbamate, 0.1 M sodium thioglycollate, 20% (v/v) carbon tetrachloride and 20% (v/v) chloroform. Centrifuge for 10 min at 8000 g and discard the sediment and solvent phase. Add 6% w/v polyethylene glycol (M. Wt 6000) to the supernatant fluid, stir the mixture for 1 h, collect the precipitate by centrifugation (15 min at 10,000 g) and resuspend for 1 h in 0. 1 M Tris buffer at pH 9.0 (1.5 ml/1 g leaf tissue). Clarify by low speed centrifugation, centrifuge supernatant fluid for 1.5 h at 55,000 g, and resuspend sedimented virus in 0.1 M Tris buffer at pH 8. The virus is further purified by density gradient centrifugation in caesium chloride (16 h at 110,000 g), reconcentrated by ultracentrifugation (1.5 h at 55,000 g), and resuspended (1 ml/100 g leaf tissue) in 0.1 M Tris buffer at pH 8.0 (Van der Meer et al., 1980).
Properties of Particles
The virus sediments as a single component.
Sedimentation coefficient (s°20, w): 157 S.
A260/A280: 1.12.
Buoyant density in caesium chloride: 1.31 g cm-3.
Particle Structure
The virus has straight or slightly flexuous filamentous particles measuring 13 x 650 nm (Fig. 6).
Particle Composition
Nucleic acid: Single-stranded RNA; one species, of M. Wt 2.8 x 106 (estimated by polyacrylamide gel electrophoresis in non-denaturing conditions), and comprising c. 5% of particle weight (estimated from the A260:A280 ratio; Brunt et al., 1980).
Protein: Single polypeptide species of M. Wt c. 31,000 (Brunt et al., 1980).
Relations with Cells and Tissues
No information.
Notes
Although honeysuckle latent virus is serologically related to poplar mosaic and shallot latent viruses, its relationship to both is sufficiently distant (serological differentiation indices of 4-8) for it to be recognised as a distinct carlavirus. The relationship between honeysuckle latent and poplar mosaic virus is particularly interesting because, as the latter has no known aphid vector and was previously considered to be serologically distinct from carlaviruses, its taxonomic status was hitherto uncertain. Its serological relationship to honeysuckle latent virus thus supports physico-chemical (Luisoni, Boccardo & Milne, 1976) and cytological (Brunt, Stace-Smith & Leung, 1976) evidence for its inclusion in the carlavirus group.
Plants of many honeysuckle cultivars can be obtained free of honeysuckle
latent virus by rooting small shoots (c. 1 cm long) from infected
plants which have been grown for 6-8 wk at alternating temperatures (16 h
at 37°C, 8 h at 26°C) (Van der Meer et al., 1980). Such
virus-free material is now commercially available in the Netherlands
(Elzenga, 1974).
Unlike honeysuckle latent virus, cucumber mosaic virus induces severe
leaf chlorosis in naturally infected honeysuckle (Lihnell, 1951; Brunt &
Thomas, 1976), and tobacco leaf curl virus causes yellow vein-banding in
Lonicera japonica var. aureoreticulata (Osaki et al.,
1979). Honeysuckle latent and cucumber mosaic viruses are transmissible
by aphids and by mechanical inoculation of sap, whereas tobacco leaf curl
virus is transmissible by whiteflies (Bemisia tabaci) but not by
mechanical inoculation. Honeysuckle latent and cucumber mosaic viruses can
be differentiated by the size, shape and properties of their particles and
by the symptoms they induce in differential hosts.
Figures

Chlorotic lesions in inoculated Chenopodium quinoa leaf.

Chlorotic spotting in systemically infected C. quinoa leaf.
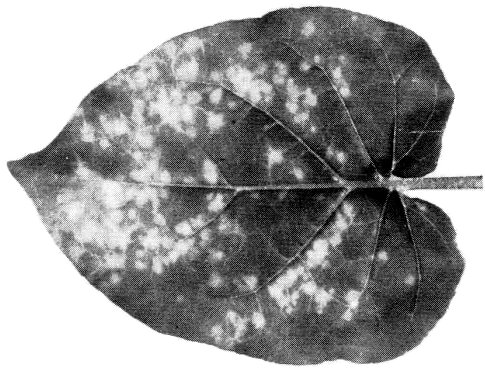
Chlorotic lesions in inoculated Nicotiana glutinosa leaf.

Faint chlorotic lesions in inoculated N. megalosiphon leaf.

Veinal chlorosis in systemically infected N. megalosiphon leaf.

Virus particle mounted in uranyl acetate. Bar represents 100 nm.(Micrograph courtesy of Dr R. G. Milne.)
References list for DPV: Honeysuckle latent virus (289)
- Bos, Huttinga & Maat, Neth. J. Pl. Path. 84: 227, 1978.
- Brunt & Thomas, Rep. Glasshouse Crops Res. Inst. for 1975, p. 124, 1976.
- Brunt, Phillips & Thomas, Acta Hort. 110: 205, 1980.
- Brunt, Stace-Smith & Leung, Intervirology 7: 303, 1976.
- Lihnell, Vaxtskyddsnotiser 15: 52, 1951.
- Luisoni, Boccardo & Milne, Phytopath. Z. 85: 65, 1976.
- Osaki, Kobatake & Inouye, Ann. phytopath. Soc. Japan 45: 62, 1979.
- Van der Meer, Rep. Res. Inst. Pl. Prot. for 1978, p. 31, 1979.
- Van der Meer, Maat & Vink, Neth. J. Pl. Path. 86: 69, 1980.