Details of DPV and References
DPV NO: 29 October 1970
Family: Secoviridae
Genus: Comovirus
Species: Broad bean stain virus | Acronym: BBSV
Broad bean stain virus
A. J. Gibbs John Curtin School of Medical Research, Australian National University, Canberra, Australia
Helen G. Smith Little Paddock, Kirklington, Newark, Nottinghamshire, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Lloyd, Smith & Jones (1965) and
Gibbs, Giussani-Belli & Smith (1968).
Synonym:
- Broad bean Evesham stain virus (Rev. appl. Mycol. 44: 3213)
-
A virus with angular isometric RNA-containing particles, about 25 nm in diameter, which sediment as three components when centrifuged. It is readily transmitted by inoculation of sap and is seed-borne, but no specific vector has been found. It seems to infect only legumes, and occurs in Europe and North Africa.
Main Diseases
Infects broad bean plants (Vicia faba) causing chlorotic mottling and mosaic of the leaves (Fig. 1, Fig. 2), and necrotic staining of the seed testa (Fig. 3).
Geographical Distribution
Europe and North-West Africa.
Host Range and Symptomatology
Hosts restricted to Leguminosae; Gibbs et al. (1968) inoculated 44 species in 19 dicotyledonous families, but the virus infected only 7 species of legume: Crotalaria spectabilis, Lupinus hirsutus, Melilotus alba, Phaseolus vulgaris (French bean), Pisum sativum (pea), Trifolium incarnatum (crimson clover) and Vicia faba (broad bean).
-
Diagnostic species
- Phaseolus vulgaris.
Varieties Tendergreen and Canadian Wonder show chlorotic local lesions, and a systemic chlorotic mosaic. The Prince is infected only locally, and Pinto, Idaho Refugee, Blue Lake and Tendercrop not at all. - Pisum sativum. All varieties tested are susceptible, and develop a
systemic chlorotic mottle, with stem and leaf necrosis in cool weather.
- Chenopodium amaranticolor, Gomphrena globosa, Nicotiana tabacum (tobacco) and N. clevelandii are not susceptible. >
-
Propagation species
- Pisum sativum
Onward, Phaseolus vulgaris cv. Tendergreen and Vicia faba.Assay species
- Phaseolus vulgaris
cv. Tendergreen, in which the virus gives local lesions, and Vicia faba, which is infected systemically.
Strains
None reported.
Transmission by Vectors
Vector not known, though the virus spreads over large distances within crops (Gibbs et al., 1968).
Transmission through Seed
Common in certain varieties of broad bean, transmitted to up to 10% of the progeny of infected plants.
Transmission by Dodder
Not tested.
Serology
Moderately immunogenic. Gives a single band of precipitate in Ouchterlony gel diffusion tests, and a granular precipitate in tube tests.
Relationships
The physical and chemical properties of the particles of this virus resemble those of cowpea mosaic virus. It is distantly serologically related to red clover mottle virus, F1 virus, and cowpea mosaic virus (Devergne, 1965, 1966; Gibbs et al., 1968; Valenta & Gressnerova, 1966).
Stability in Sap
In broad bean sap the thermal inactivation point (10 min) is between 60° and 65°C. Infectivity survives at least 31 days at 4°C and more than one year at -15°C. The dilution end-point in sap from leaves showing obvious symptoms is somewhat variable but usually around 10-3.
Purification
The virus particles are stable and Steere’s butanol/chloroform method may be used to extract them from plant tissue into a pH 7.0 buffer containing a reducing agent. They may then be sedimented and purified by differential centrifugation (Gibbs et al., 1968).
Properties of Particles
The particles are all the same size but sediment as three components (Fig. 4). Sedimentation coefficients (s20,w) are about 60, 100 and 127 S.
Particle Structure
Particles are isometric, about 25 nm in diameter with angular polygonal outlines (Fig. 5). The 60 S particles are penetrated by negative stain, and then show a central cavity with a surrounding shell 2.0-2.5 nm thick. The 100 S particles are partially penetrated by stain, and the 127 S particles not at all. The particles show no regularly arranged morphological subunits.
Particle Composition
RNA: Probably single-stranded. Molar percentages of nucleotides: G23.3; A26.5; C18.4; U31.9. Their sedimentation coefficients suggest that the 127 S particles contain about 35% RNA, and the 100 S particles about 25%; the 60 S particles contain none.
Protein. The size and composition of the protein subunits are not yet known.
Relations with Cells and Tissues
No information.
Notes
Readily confused with Echtes Ackerbohnemosaik-Virus, because both viruses cause similar leaf symptoms in broad bean, are seed-borne, and have similar particles. However, Echtes Ackerbohnemosaic-Virus rarely infects Phaseolus vulgaris, is serologically unrelated to broad bean stain virus, and preparations of its particles contain few or no (60 S) RNA-free particles (Fig. 5).
Acknowledgements
Photographs: courtesy of Rothamsted Experimental Station, UK.
Figures

Broad bean (Vicia faba) plant infected with broad bean stain virus through the seed.

Two systemically infected broad bean leaves.

Seeds of infected broad beans showing the characteristic brown necrosis of the testa around the periphery of the seed.

Sedimentation behaviour of the virus (upper schlieren diagram) compared with that of Echtes Ackerbohnemosaik-Virus (lower diagram); sedimentation is from left to right (note that the menisci of the two diagrams are displaced).
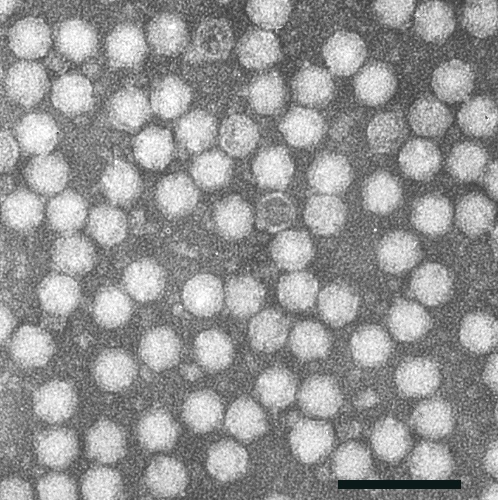
Electron micrograph of a phosphotungstate-stained purified preparation of the virus. Bar represents 100 nm.
References list for DPV: Broad bean stain virus (29)
- Devergne, Annls Épiphyt. 16: 41, 1965.
- Devergne, Annls Épiphyt. 17: 147, 1966.
- Gibbs, Giussani-Belli & Smith, Ann. appl. Biol. 61: 99, 1968.
- Lloyd, Smith & Jones, Hort. Res. 5: 13, 1965.
- Valenta & Gressnerova, Acta virol., Prague 10: 182, 1966.