Details of DPV and References
DPV NO: 3 June 1970
Family: Bromoviridae
Genus: Bromovirus
Species: Brome mosaic virus | Acronym: BMV
There is a more recent description of this virus: DPV 180
Brome mosaic virus
J. B. Bancroft Purdue University, Lafayette, Indiana, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
McKinney, Fellows & Johnston (1942).
Selected synonyms
- Weidelgrasmosaik-Virus (Rev. appl. Mycol. 39: 589)
- Ryegrass streak virus (Rev. appl. Mycol. 44: 152)
- Trespenmosaik-Virus (Rev. appl. Mycol. 44: 2517)
- Marmor gramminis (Rev. appl. Mycol. 23: 427)
- Ryegrass streak virus (Rev. appl. Mycol. 44: 152)
-
An RNA-containing virus with isometric particles about 25 nm in diameter. It is readily transmissible by inoculation with sap and infects many monocotyledonous plants and a few dicotyledonous ones. It is found in temperate regions and is reported to be transmitted by nematodes (Xiphinema spp.).
Main Diseases
Causes a usually mild mosaic in most species of the Gramineae.
Geographical Distribution
Reported from the USA and Europe.
Host Range and Symptomatology
Monocotyledonous host range is large, comprising about 60 genera in the Gramineae; dicotyledonous host range is restricted to a few genera in about 5 families (McKinney, 1944; Ohmann-Kreutzberg, 1963).
-
Diagnostic species
- Zea mays
(maize). Seedlings of most varieties of sweet corn develop primary lesions or streaks (Fig. 1) followed by systemic necrosis and death (Fig. 2).Propagation species
- Hordeum vulgare
(barley). Mild mosaic.Assay species
- Chenopodium hybridum
(Rochow, 1959) (Fig. 3) and Datura stramonium (Chiu & Sill, 1963) are local lesion hosts.
Strains
Although the virus has been isolated from many locations and the existence of variants implied, no direct systematic comparisons have been published. In the USA, the McKinney isolate (McKinney et al., 1942) is regarded as the type strain as opposed to the Sill isolate (Chiu & Sill, 1963) which is usually assumed to be different.
Transmission by Vectors
The only positive transmission trials have been those reported by Schmidt, Fritzsche & Lehmann (1963) who claim Xiphinema paraelongatum (X. diversicaudatum) and another species, subsequently identified as X. coxi, as vectors.
Transmission through Seed
None reported.
Transmission by Dodder
No information.
Serology
The virus is moderately immunogenic (Moorhead, 1956). It produces single bands in gel-diffusion tests at pH 5 but may produce 2 or 3 bands at neutrality or higher (Hamilton, 1961).
Relationships
All isolates of the virus cross react serologically. Quantitative differences have not been reported, but may occur.
Stability in Sap
The thermal inactivation point (10 min) in barley sap is about 79°C, and the dilution end-point is between 1 x 10-5 and 3 x 10-5. The virus survives in air-dried leaf tissue for more than 1 year.
Purification
The virus, which occurs in concentrations of about 1 g/l of barley juice, is extremely easy to purify. A convenient schedule, based on that of Bockstahler & Kaesberg (1962), is to blend tissue, collected about 2 weeks after inoculation, in 0.2 M pH 4.8 acetate buffer, express the homogenate through cheesecloth, allow it to stand for a few hours to overnight at 4°C, and then further purify and concentrate by 2 to 3 cycles of differential ultracentrifugation. Resuspend pellets in 0.1 M pH 5.0 acetate buffer and store at 4°C.
Properties of Particles
Sedimentation coefficient (s20, w): (87.3-0.47c) S at pH 3-6 and (78.7-0.64c) S at pH 7 and above (Incardona & Kaesberg, 1964) where c = nucleoprotein concentration in mg/ml. The virus sediments as a single component at pH 3-6 but may undergo degradation and sediment as two or more components at pH 7 or above depending on dialysis time and exact ionic conditions. The virus is inactivated at pH 7.
Molecular weight (daltons): 4.6 x 106 (Bockstahler & Kaesberg, 1962).
Isoelectric point: about pH 7.9 at ionic strength of 0.1 (type strain).
Electrophoretic mobility: +1.41 x 10-5 cm2 sec-1 volt-1 at pH 6 at an ionic strength of 0.1 (type strain).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 5.2 with no scattering correction.
A260/A280: 1.7.
Buoyant density: There are two density populations in CsCl at close to the average value of 1.363 g/ml. The less dense population probably results from a terminal RNA deletion of about 250 nucleotides and is probably not infective.
Particle Structure
Particles are isometric (Fig. 4), about 250 nm in diameter in 1% pH 4.7 uranyl acetate, and are built of 180 structure units in hexamer-pentamer clusters (Bancroft, Hills & Markham, 1967). The particles have a small, sometimes noticeable, electron-dense central area about 5 to 7.5 nm in diameter.
Particle Composition
RNA: Molecular weight is 1 x 106; presumably derivative RNA molecules of 0.3 x 106 and 0.7 x 106 occur together in some particles. Single-stranded. Molar percentages of nucleotides: G28; A27; C21; U24. RNA is 21-22% of particle weight. Sedimentation coefficients (s20, w) in 0.1 M KCl + 2 x 10-3 M divalent cations at pH 5.5 are 26.8, 22.3 and 14 S for the 3 major classes of RNA (Bockstahler & Kaesberg, 1965).
Protein: Subunits have a molecular weight of about 20,300 and contain 189 amino acid residues (Stubbs & Kaesberg, 1964).
Relations with Cells and Tissues
No information.
Notes
The virus is physically similar to cowpea chlorotic mottle and broad bean mottle viruses but is serologically related to neither. Neither of these viruses infects barley or sweet corn.
Figures
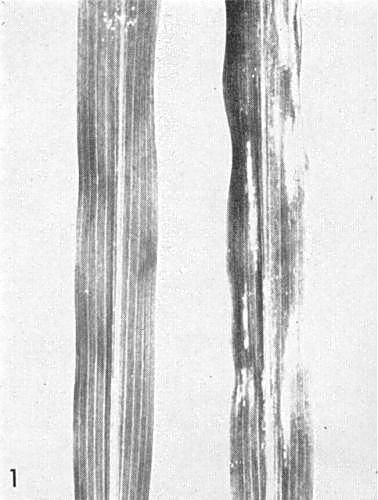
Sweet corn leaves, (left) healthy, (right) infected, 5 days after inoculation.

Sweet corn plants, (left) healthy, (right) infected, 10 days after inoculation.

Local lesions in Chenopodium hybridum.

Virus particles from a purified preparation in 1% pH 4.7 uranyl acetate. Bar represents 100 nm. Enlarged particle in inset shows detailed structure.
References list for DPV: Brome mosaic virus (3)
- Bancroft, Hills & Markham, Virology 31: 354, 1967.
- Bockstahler & Kaesberg, Biophys. J. 2: 1, 1962.
- Bockstahler & Kaesberg, J. molec. Biol. 13: 127, 1965.
- Chiu & Sill, Phytopathology 53: 69, 1963.
- Hamilton, Virology 15: 452, 1961.
- Incardona & Kaesberg, Biophys. J. 4: 11, 1964.
- McKinney, Phytopathology 34: 993, 1944.
- McKinney, Fellows & Johnston, Phytopathology 32: 331, 1942.
- Moorhead, Phytopathology 46: 498, 1956.
- Ohmann-Kreutzberg, Phytopath. Z. 47: 1, 1963.
- Rochow, Phytopathology 49: 126, 1959.
- Schmidt, Fritzsche & Lehmann, Naturwissenschaften 50: 386, 1963.
- Stubbs & Kaesberg, J. molec. Biol. 8: 314, 1964.