Details of DPV and References
DPV NO: 307 September 1985
Family: Bromoviridae
Genus: Ilarvirus
Species: Tobacco streak virus | Acronym: TSV
There is a more recent description of this virus: DPV 381
This is a revised version of DPV 44
Tobacco streak virus
R. W. Fulton Department of Plant Pathology, University of Wisconsin, Madison, Wisconsin, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Johnson (1936).
Selected synonyms
- Annulus orae
(Rev. appl. Mycol. 28: 514) - Asparagus stunt virus (Rev. Pl. Path. 50, 1046h)
- Datura ‘quercina’ virus (Rev. appl. Mycol. 1: 74)
- Nicotiana virus 8 (Rev. appl. Mycol. 36: 303)
- Nicotianavirus vulnerans (Rev. appl. Mycol. 36: 303)
- Datura ‘quercina’ virus (Rev. appl. Mycol. 1: 74)
-
A virus with isometric RNA-containing particles 27 to 35 nm in diameter. It has a wide host range. It is readily transmitted by manual inoculation, but is unstable in plant extracts. There are two reports of transmission by thrips. The virus is widespread, but does not often occur in epidemic proportions.
Main Diseases
Causes tan systemic necrotic lines bordering veins (streak, 'necrose branca') in tobacco (Fig. 1), with subsequent recovery from symptoms; mottling or no symptoms in dahlia (Costa & Carvalho, 1961; Brunt, 1968); mottling in cotton (Costa & Carvalho, 1961), Trifolium pratense and Melilotus alba; yellow ringspot and malformation in tomato (Martelli & Cirulli, 1969; Costa et al., 1961); stunting of asparagus (Brunt & Paludan, 1970); vein yellowing in rose (Fulton, 1970b); red node in bean (Thomas & Zaumeyer, 1950); systemic necrosis in pea (Patino & Zaumeyer, 1959), soybean (Costa & Carvalho, 1961; Fagbenle & Ford, 1970) and potato (Costa et al., 1964); and necrotic shock in strawberry (Stace-Smith & Frazier, 1971). It has also been isolated from grape (J. K. Uyemoto, personal communication), papaya (A. F. Ross, personal communication), black raspberry (Converse, 1972), red raspberry (Stace-Smith et al., 1982), blackberry (Jones & Mayo, 1975), groundnut, pepper (Gracia & Feldman, 1974), globe artichoke (Costa & Tasaka, 1971), alfalfa (Paliwal, 1982), sunflower (Dijkstra, 1983) and numerous wild species.
Geographical Distribution
Europe, North and South America, Japan, Australia and New Zealand.
Host Range and Symptomatology
The host range is wide; many species in over 30 monocotyledonous and dicotyledonous families are susceptible.
-
Diagnostic species
- Nicotiana tabacum
(tobacco). Local necrotic spots or rings, systemic necrotic lines and ‘oak leaf’ patterns; plants recover from necrotic symptoms, and leaves that expand several weeks after infection may appear healthy (Fig. 1). With many strains of the virus, ‘recovered’ leaves of Turkish tobacco have dentate rather than entire margins (Fig. 2) (Berkeley & Phillips, 1943) and petal terminations are filamentous (Costa, 1945). Isolates from Rubus and dahlia infect tobacco with difficulty or not at all. - Cyamopsis tetragonoloba (guar). Small, dark local lesions
(Fig. 4).
- Vigna unguiculata ssp. cylindrica (catjang). Strains may give either local reddish necrotic lesions or chlorotic lesions (Fig. 5, Fig. 6) with subsequent systemic necrosis or mottle.
-
Propagation species
- Cultures of most isolates are readily maintained in tobacco or Vinca rosea.
Tobacco, Nicotiana rustica, Datura stramonium and Vigna unguiculata
ssp. cylindrica are good sources of virus for purification. D. stramonium
has yielded several times as much virus of many strains as other species
(Fulton & Potter, 1971).
Isolates from Rubus are best propagated in Chenopodium
quinoa.
Assay species
- V. unguiculata
ssp. cylindrica, Dolichos biflorus, Beta patellaris, Phaseolus vulgaris cv. Manteiga and Gomphrena globosa have been used as local lesion hosts with various isolates.
Strains
Tobacco streak virus exists in a variety of strains. Several isolates from South America differed serologically from common North American isolates (Fulton, 1972; Ghabrial & Lister, 1974) as did isolates from North Carolina (Gooding, 1971), Washington state (Kaiser et al., 1982) and from black raspberry (Jones & Mayo, 1975). Isolates may also differ in symptoms caused on indicator hosts. Isolates from Rubus (Converse, 1972; Jones & Mayo, 1975) and dahlia (R. W. Fulton, unpublished data) can be transmitted to tobacco only with difficulty or not at all. Black raspberry latent virus (Converse & Lister, 1969) is considered a distinct strain (Jones & Mayo, 1975). Bean red node virus (Thomas & Zaumeyer, 1950) is probably also a distinct strain.
Transmission by Vectors
A Frankliniella sp. has been reported as a vector in Brazil by Costa & da Costa Lima Neto (1976). A mixture of Thrips tabaci and F. occidentalis was described as transmitting the virus in USA (Kaiser et al., 1982).
Transmission through Seed
Reported for bean, Datura stramonium, Chenopodium quinoa (Brunt, 1969), Melilotus alba, Glycine max, Gomphrena globosa, Nicotiana clevelandii, Vigna unguiculata (Kaiser et al., 1982), black raspberry (Converse & Lister, 1969) and Nicandra physalodes (Salazar et al., 1982). Reported frequencies of transmission range from 90% in Glycine max to less than 1% in Vigna unguiculata (Kaiser et al., 1982). The black raspberry latent strain is transmitted by pollen not only to seed but also to the plant pollinated (Converse & Lister, 1969).
Transmission by Dodder
Transmitted by Cuscuta campestris (Fulton, 1948).
Serology
The virus is moderately immunogenic; 12-14 intramuscular injections of 1-2 mg of virus in Freund’s incomplete adjuvant at 3-4 day intervals induce antiserum titres of 1/2560-1/5120 (by microprecipitin test). Precipitates in liquid tests are granular. The virus gives a well defined zone of precipitate in agar gels made with saline. Strain-specific antisera have been prepared by cross absorption with purified particles of various virus strains.
Relationships
All strains cross react with each other’s antisera, but some have strain-specific antigenic determinants. Most strains reciprocally protect against each other in plants, but pairs of strains that protect only unilaterally, or not at all, have been described (Fulton, 1978). Inability to cross-protect is not correlated with serological difference. The virus was designated as the type member of the ilarvirus group (Shepherd et al., 1975/1976), which is composed of viruses having three particle types of different sizes, but each with the same (14%) RNA content. Van Vloten-Doting et al. (1981) proposed that the ilarvirus group be considered a genus in a family consisting also of cucumoviruses and bromoviruses.
Stability in Sap
Undiluted tobacco sap loses most of its infectivity within 5 min after extraction (Fulton, 1949) and all of its infectivity in less than 36 h. Infectivity is lost more slowly in diluted extracts and is stabilized by antioxidants, especially 2-mercaptoethanol. Thermal inactivation points between 53 and 64°C and dilution end-points between 1/30 and 1/15,625 are reported; variation is apparently caused by the presence or absence of antioxidants. The virus remains infective for many years in diced tissue dried and stored at about 2°C.
Purification
The following method is effective (Fulton, 1967). Homogenize heavily infected leaf tissue about 10 days after local manual inoculation at the rate of 100 g in 150 ml buffer (0.02 M phosphate, pH 8.0, containing 0.02 M 2-mercaptoethanol) plus 15 g Al2O3. After centrifuging 15-20 min at 1500 g, mix the supernatant liquid thoroughly with 80 ml hydrated calcium phosphate per 100 g tissue. Centrifuge again at 1500 g for 15-20 min. Sediment the virus from the supernatant liquid by centrifuging 3 h at 78,000 g. Resuspend the pellets in 0.01 M disodium ethylene diamine tetraacetate (EDTA), pH 6.0, adjust the pH to 5.0 with citric acid and remove the precipitate by centrifugation. Readjust the supernatant liquid, which contains the virus, to pH 6.0 with NaOH and concentrate the virus by high speed centrifugation. Resuspend the pellets in 0.01 M EDTA, pH 6.0, in which the virus is more stable than in water or a number of buffers (Fulton, 1970a). Yields of virus from infected Datura stramonium may be up to 400 µg per gram of tissue (Fulton & Potter, 1971). Certain isolates of the virus cannot be purified by this method and require a method involving precipitation with 4% polyethylene glycol, M. Wt 6000, and 0.1 M NaCl (Fulton, 1978). Procedures involving clarification of leaf extracts by charcoal and freezing (Mink et al., 1966) or by the ether-CCl4 method of Wetter (Greber, 1971) have also been used.
Properties of Particles
The virus has three main kinds of nucleoprotein particle (Fig. 3). Sedimentation coefficients (s20, w) are c. 90, 98, and 113 S (Lister & Bancroft, 1970). The relative proportions of the particle types may vary with the host, purification procedure (Lister & Bancroft, 1970), strain, or time after inoculation (Fulton & Potter, 1971). Infectivity of separated particle types is nil or nearly so for top (T) and middle (M) particles, and low for bottom (B) particles (Fulton, 1970a). Maximum infectivity is obtained with mixtures of M and B particles. Supplementing such mixtures with T particles increases infectivity, but increasing the proportion of T particles in mixtures of constant concentration decreases infectivity (Fulton, 1975). T, M and B particles have M. Wts (x 10-6) of 4.72, 5.92 and 7.45 (Ghabrial & Lister, 1974).
Diffusion coefficients (x 10-7 cm2/sec) of T, M and B particles are 1.39, 1.32 and 1.23.
The isoelectric point of two strains was pH 4.6 in 0.1 ionic strength buffer.
Partial specific volume: 0.71 ml/g.
Absorbance at 260 nm (1 µg/ml, 1 cm light path): 5.1.
A260/A280: c. 1.56 for each particle type.
Buoyant density in CsCl: 1.35 g/cm3 for each particle type (Jones & Mayo, 1975).
Particle Structure
Particles are isometric, of three major sizes: 27, 30 and 35 nm in diameter (Fig. 3) (Lister et al., 1972). The numbers of protein subunits for the three particle types are thought to be: T, 142; M, 179; and B, 225 (Ghabrial & Lister, 1974). Regularity in arrangement of subunits is not apparent in electron micrographs, which show slightly distorted particles without hollow centres (Fig. 3).
Particle Composition
Nucleic acid: Single-stranded RNA, divided into three major genome segments, with M. Wt (x 106) of 1.35, 1.1 and 0.85; one subgenomic RNA of M. Wt 0.4 x 106 is also found in the virus particles (Jones & Mayo, 1975). The RNA comprises c. 14% of each particle type (Ghabrial & Lister, 1974). The molar percentages of nucleotides (G:A:C:U) are 25:25:20:30. The RNA molecules do not accept amino acids, but have high affinity sites for coat protein (Van Vloten-Doting, 1975).
Protein: Comprises 86% of the particle weight. The M. Wt of the protein subunits is 28,000. The amino acid composition of two serologically different strains differed, especially in contents of alanine, aspartic acid, leucine, proline and valine (Ghabrial & Lister, 1974).
Genome Properties
Mixtures of the three main RNA species are not infective, but become so in the presence of the smallest RNA or of coat protein. Coat protein of alfalfa mosaic virus, although unrelated serologically to that of tobacco streak virus, also activates infectivity. Similarly, coat protein of tobacco streak virus activates the three largest RNA molecules of alfalfa mosaic virus (Van Vloten-Doting, 1975). Tobacco streak virus protein also activated nucleic acid mixtures of citrus leaf rugose and citrus variegation viruses, ilarviruses serologically unrelated to tobacco streak virus (Gonsalves & Garnsey, 1975). Van Vloten-Doting (1975) reported that the largest genomic RNA molecule occurs in B virus particles, the intermediate RNA molecule in M virus particles, and the smallest RNA molecule in T virus particles. Heterogeneity in RNA content of M particles has been described, however (Clark & Lister 1971), and this agrees with genetic data indicating that determinants for lesion type in Vigna unguiculata ssp. cylindrica may be located in either T or M particles (Fulton, 1970a). By analogy with alfalfa mosaic virus, the gene for the coat protein is probably contained in the 0.85 x 106 M. Wt RNA and presumably also in the 0.4 x 106 M. Wt RNA, which is probably a subgenomic fragment of the 0.85 x 106 M. Wt RNA. Genetic evidence, obtained by inoculating heterologous mixtures of particle types and cloning pseudo-recombinant isolates from single lesions, indicated that serological type might correspond to the source of either M or B particles. A severe dwarfing symptom in Turkish tobacco (Fig. 7) is conditioned by two genes, one contained in M and the other in B particles (Fulton, 1972). Size and colour of lesions in Vigna are inherited independently, the gene for small lesions being frequently carried in T particles (Fulton, 1975). An unstable gene conditioning a ‘non-recovering’ symptom in tobacco was carried only in T particles.
Relations with Cells and Tissues
Virus particles occur in aggregates in the cytoplasm and nucleus of hosts (Edwardson & Purcifull, 1974). The virus was obtained from embryos and coats of immature soybean seeds, but only from the embryos of mature seeds (Ghanekar & Schwenk, 1980). In cotton the virus does not become systemic unless ‘anthocyanosis virus’ is also present (Costa, 1969).
Notes
Symptoms of the virus in diagnostic hosts may vary and may be confused with those of other viruses. Identification should be made by serology. In some instances single isolates of the virus have been described as strains without evidence that they differ significantly from other isolates. Some of the data in this description were obtained with only one or two isolates and may not be representative of all isolates.
Figures

Systemic necrotic symptoms and ‘recovery’ in tobacco cv. Xanthi-nc.

Serrations in ‘recovered’ leaves of Xanthi-nc tobacco.
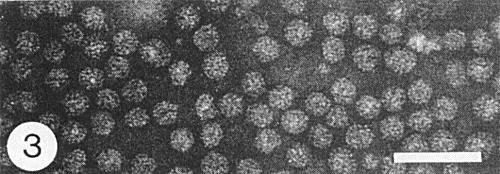
Purified virus particles, fixed in 1% glutaraldehyde and mounted in phosphotungstate. Bar represents 100 nm.

Local necrotic lesions in Cyamopsis tetragonoloba.

Local lesions in Vigna unguiculata ssp. cylindrica. Large necrotic lesions of strain WC (left) and small necrotic lesions of strain HF (right).

Local lesions in Vigna unguiculata ssp. cylindrica. Chlorotic lesions of strain B.

Dwarfing in Xanthi-nc tobacco resulting from two mutant genes of the virus, one carried in M, the other in B particles.
References list for DPV: Tobacco streak virus (307)
- Berkeley & Phillips, Can. J. Res. 21: 181, 1943.
- Brunt, Pl. Path. 17: 119, 1968.
- Brunt & Paludan, Phytopath. Z. 69: 277, 1970.
- Clark & Lister, Virology 45: 61, 1971.
- Converse, Phytopathology 62: 1001, 1972.
- Converse & Lister, Phytopathology 59: 325, 1969.
- Costa, Phytopathology 35: 1029, 1945.
- Costa, Phytopath. Z. 65: 219, 1969.
- Costa & Carvalho, Phytopath. Z. 43: 113, 1961.
- Costa & da Costa Lima Neto, Proc. 9th Cong. Soc. Bras. Fitopath.1 p., 1976.
- Costa & Tasaka, Biologico 37: 176, 1971.
- Costa, Carvalho, Oliveira & Deslandes, Bragantia 20: cvii, 1961.
- Costa, Carvalho & Deslandes, Bragantia 23: 1, 1964.
- Dijkstra, Neth. J. Pl. Path. 89: 153, 1983.
- Edwardson & Purcifull, Phytopathology 64: 1322, 1974.
- Fagbenle & Ford, Phytopathology 60: 814, 1970.
- Fulton, Phytopathology 38: 421, 1948.
- Fulton, Phytopathology 39: 231, 1949.
- Fulton, Virology 32: 153, 1967.
- Fulton, Virology 41: 288, 1970a.
- Fulton, Pl. Dis. Reptr 54: 949, 1970b.
- Fulton, Virology 50: 810, 1972.
- Fulton, Virology 67: 188, 1975.
- Fulton, Virology 85: 1, 1978.
- Fulton & Potter, Virology 45: 734, 1971.
- Ghabrial & Lister, Virology 57: 1, 1974.
- Ghanekar & Schwenk, Phytopath. Z. 97: 148, 1980.
- Gonsalves & Garnsey, Virology 67: 319, 1975.
- Gooding, Phytopathology 61: 1303, 1971.
- Gracia & Feldman, Phytopath. Z. 80: 313, 1974.
- Greber, Qd. J. agric. Anim. Sci. 28: 105, 1971.
- Johnson, Phytopathology 26: 285, 1936.
- Jones & Mayo, Ann. appl. Biol. 79: 297, 1975.
- Kaiser, Wyatt & Pesho, Phytopathology 72: 1508, 1982.
- Lister & Bancroft, Phytopathology 60: 689, 1970.
- Lister, Ghabrial & Saksena, Virology 49: 290, 1972.
- Martelli & Cirulli, Phytopath. Mediterranea 8: 154, 1969.
- Mink, Saksena & Silbernagel, Phytopathology 56: 645, 1966.
- Paliwal, Can. J. Pl. Path. 4: 175, 1982.
- Patino & Zaumeyer, Phytopathology 49: 43, 1959.
- Salazar, Abad & Hooker, Phytopathology 72: 1550, 1982.
- Shepherd, Francki, Hirth, Hollings, Inouye, Macleod, Purcifull, Sinha, Tremaine,Valenta & Wetter, Intervirology 6:181, 1975/1976.
- Stace-Smith & Frazier, Phytopathology 61: 757, 1971.
- Stace-Smith, Daubeny, Bristow & Baumann, Acta Hort. 129: 91, 1982.
- Thomas & Zaumeyer, Phytopathology 40: 832, 1950.
- Van Vloten-Doting, Virology 65: 215, 1975.
- Van Vloten-Doting, Francki, Fulton, Kaper & Lane, Intervirology 15: 198, 1981.