Details of DPV and References
DPV NO: 31 October 1970
Family: Betaflexiviridae
Genus: Capillovirus
Species: Apple stem grooving virus | Acronym: ASGV
There is a more recent description of this virus: DPV 376
Apple stem grooving virus
R. M. Lister Purdue University, Lafayette, Indiana, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Lister, Bancroft & Nadakavukaren (1965),
de Sequeira (1965;
1967)
and
de Sequeira & Lister (1969b).
Selected synonyms
-
Apple latent virus Type 2 (C-431 isolate) (Rev. appl. Mycol. 45: 134)
- E-36 virus (Rev. appl. Mycol. 45: 3362d; 46: 3488)
- Virginia Crab stem grooving virus (de Sequeira & Cropley, 1968)
- Dark green epinasty virus (in Chenopodium quinoa) (Rev. appl. Mycol. 48: 2454)
- Brown line virus disease (Mink et al., 1970)
- E-36 virus (Rev. appl. Mycol. 45: 3362d; 46: 3488)
-
A virus with very flexuous filamentous particles c. 600-700 x 12 nm. Known only from apple (Malus sylvestris) from which it is sap-transmissible to some herbaceous species. Its natural mode of spread is unknown, but it occurs symptomlessly in many commercial apple clones.
Main Diseases
Causes stem grooving, brown line and graft union abnormalities in Virginia Crab, the only apple cultivar definitely known to show symptoms.
Geographical Distribution
Reported in apple clones of N. American, English, Dutch, Portuguese, Italian, Japanese and Indian provenance. Probably occurs wherever apples are cultivated.
Host Range and Symptomatology
Host range moderately wide. Reported to infect about 20 species in 9 dicotyledonous families (Aizoaceae, Amaranthaceae, Chenopodiaceae, Cucurbitaceae, Labiatae, Leguminosae, Rosaceae, Scrophulariaceae, Solanaceae), many symptomlessly (Lister et al., 1965; de Sequeira, 1967; Waterworth & Gilmer, 1969). Transmissible from apple, especially in spring, by inoculating extracts from buds, young leaves or petals ground in 0.05 M phosphate buffer at pH 7-8. Additives such as 2% (v/v) nicotine base, or 2% (w/v) polyvinyl pyrrolidone, or hide powder help but are not essential. Such primary inoculations usually induce only a few local lesions or slowly-developing systemic symptoms, and subculturing is essential for bulk propagation of virus.
-
Diagnostic species
- Malus sylvestris
cv. Virginia Crab (apple). Elongated grooves in the wood, visible on removal of bark (de Sequeira, 1967; Lister, Bancroft & Shay, 1967) (Fig. 7). The ‘E-36’ strain also causes swelling at the union of the Virginia Crab scion with the rootstock, flattening or sunken areas on the stem, and browning and fragility at the graft union (Fig. 4, Fig. 5, Fig. 6). Some isolates cause no symptoms in Virginia Crab (Waterworth & Gilmer, 1969). (For a detailed description and differentiation from the ‘apple stem pitting disease’, also characterized in Virginia Crab, see de Sequeira & Cropley (1968), de Sequeira & Posnette (1969)). -
Chenopodium quinoa. Some isolates induce etched necrotic lesions 0.5-2 mm in
diameter in inoculated leaves
(Fig. 2).
Usual systemic symptoms are chlorotic ring and
mottle patterns, shoot tip distortion, and stunting of growth; some isolates induce
epinasty, and an abnormally deep green colour in old uninoculated leaves
(Fig. 3).
Primary lesions develop 2-3 days later than those of
apple chlorotic leaf spot virus.
When both viruses are present, systemic symptoms are much more severe than those
produced by either virus alone.
- Nicotiana glutinosa. Systemic yellow mosaic and line patterns mainly affecting young leaves, with occasional necrosis.
- Phaseolus vulgaris. Purple-brown spots or rings 0.5-3 mm in diameter, or chlorotic lesions (Fig. 1). Most varieties develop systemic veinal necrosis (de Sequeira & Lister, 1969b). Symptoms are most severe in winter.
- Nicotiana glutinosa. Systemic yellow mosaic and line patterns mainly affecting young leaves, with occasional necrosis.
-
Propagation species
- C. quinoa
is the most useful plant for bulk propagation of virus. For maintaining cultures N. glutinosa and P. vulgaris are useful in restricting other possible contaminating viruses from apple, e.g. tobacco mosaic and apple chlorotic leaf spot viruses.Assay species
- C. quinoa
and P. vulgaris cv. Pinto are useful primary-lesion hosts.
Strains
Strains studied do not differ in serological or physical properties. The following have been differentiated by symptomatology alone:
C-431 (apple latent virus type 2) (Lister et al., 1965): induces necrotic primary lesions up to 0.5 mm diameter in inoculated leaves of C. quinoa, and mild systemic mosaic and chlorotic line patterns with little stunting. It induced only stem-grooving symptoms in Virginia Crab (Lister et al., 1967).
E-36 virus (de Sequeira, 1965; 1967): no primary lesions in C. quinoa; systemic symptoms severe, with shoot tip distortion, leaf deformation and stunting. It induced stem grooving, graft union abnormalities and necrosis in Virginia Crab.
Dark green epinasty virus (Waterworth & Gilmer, 1969): several differing isolates described, most inducing in C. quinoa necrotic primary lesions 1-2 mm in diameter and systemic chlorotic ring mottle with severe stunting, shoot tip distortion and severe epinasty of non-inoculated older leaves, which became abnormally dark green. Most isolates failed to induce grooving or other abnormality in Virginia Crab.
Transmission by Vectors
No vectors known but, according to de Sequeira & Posnette (1969), spread by means other than grafting occurs occasionally.
Transmission through Seed
None reported.
Transmission by Dodder
Not transmitted by dodder in the single reported trial (Waterworth & Gilmer, 1969).
Serology
Moderately antigenic, giving antisera with titres up to 1/512. Antigenicity not detectably modified by treating the virus with glutaraldehyde before injection. Ring-precipitin tests can be used with purified virus: latex flocculation tests give specific positive results with crude buffered extracts from C. quinoa but not with extracts from apple petals or leaves (de Sequeira & Lister, 1969a, 1969b). No reactions occur in gel-diffusion tests.
Relationships
Strains examined do not differ serologically. No plant protection tests to determine strain relationships have been reported. Stem grooving virus seemed unrelated in serological and plant protection tests to the morphologically similar virus, apple chlorotic leaf spot (Lister et al., 1965; de Sequeira, 1967). In cross-protection experiments in Virginia Crab apple it seemed unrelated to an entity causing true apple stem pitting disease (de Sequeira & Cropley, 1968).
Stability in Sap
In bentonite-clarified C. quinoa sap (see below), thermal inactivation point (10 min) is between 60 and 63°C, half-life at 45°C is 105 ± 8 min, dilution end-point is about 10-4, and infectivity is retained for more than 2 days at 20°C and more than 27 days at 4°C. Reducing agents do not seem important for preserving stability.
Purification
The following methods are developments by de Sequeira & Lister (1969b) of the bentonite clarification procedure of Lister et al. (1965):
Blend each 100 g infected C. quinoa leaves in 200 ml 0.05 M phosphate buffer (pH 7-8) containing 0.15% sodium thioglycollate. Squeeze through cheesecloth and thoroughly clarify the extract by cautiously adding a bentonite suspension (c. 40 mg/ml) made in 0.01 M phosphate buffer, pH 7-8. Centrifuge at low speed. Precipitate the virus from the supernatant fluid by either (i) adjusting it to pH 4.8 and letting it stand overnight, or (ii) adding polyethylene glycol, M. Wt 6000, to 4-8% (w/v) and NaCl to 0.02 M, and letting the mixture stand 1 hr. Centrifuge at low speed and resuspend pellets in 0.05 M phosphate buffer. Concentrate by ultracentifrugation and purify further by density gradient centrifugation. Do all steps at c. 4°C. Yields are low, about 0.25 mg/100 g leaf tissue.
Partial clarification with charcoal prior to clarification with bentonite has also been tried (Waterworth & Gilmer, 1969) but the degree of purity of the product was not fully assessed.
Properties of Particles
Sedimentation coefficient (s20,w): c. 112 S at
0.3-0.6 mg/ml of 0.01 M neutral phosphate buffer. No accessory particles detected.
Isoelectric point: probably c. pH 4.3 at ionic strength of 0.1.
Electrophoretic mobility: -10.3 and -6.5 x 10-5 cm2 sec-1 volt-1 respectively at pH 7.0 and pH 6.0 (ionic strength = 0.1).
A260/A280: 1.18.
Particle Structure
Very flexuous filaments with obvious cross-banding and helical symmetry, but showing no axial canal (Fig. 8). Length 619 ± 14 nm; width c. 12 nm; pitch of helix c. 3.8 nm. Round particles, assumed to be fragments of the virus seen ‘end-on’, show a hollow centre. Best stains for electron microscopy are uranyl acetate and uranyl formate: stretching and breakage occur in phosphotungstate (Lister et al., 1965 and unpublished; de Sequeira & Lister, 1969b).
Particle Composition
Not known.
Relations with Cells and Tissues
Unknown: symptoms in Virginia Crab apple suggest an association with the cambium. Has been eliminated from some apple scions by heat therapy for 30 days at 36°C (de Sequeira & Posnette, 1969).
Notes
Specific causation of a stem grooving disease in Virginia Crab has been reported only for the E-36 and C-431 strains, respectively, by de Sequeira (1967) and Lister et al. (1967 - symptoms described as ‘stem-pitting’). Close relationship of these two strains, and of some isolates of the ‘dark green epinasty virus’ of Waterworth & Gilmer (1969), was established by de Sequeira & Lister (1969a, 1969b). Stem grooving virus is probably unrelated to the entity causing the true stem pitting disease of Virginia Crab. Because of the wide range of symptoms caused by strains of stem grooving virus both in woody and in herbaceous indicator plants, serological and/or thorough property testing is essential for accurate diagnosis.
Acknowledgements
Figures 1, 2, 7, 8 courtesy of Purdue Agricultural Experiment Station; remainder courtesy of East Malling Research Station.
Figures

Lesions caused by the ‘C-431’ strain in inoculated leaves of Phaseolus vulgaris cv. ‘Pinto’.

Lesions caused by the ‘C-431’ strain in inoculated leaves of Chenopodium quinoa.

Lesions caused by the ‘C-431’ strain in inoculated leaves of Chenopodium quinoa.

Union of 1-yr-old Virginia Crab scion with healthy rootstock.

Union of 1-yr-old Virginia Crab scion with rootstock infected with ‘E-36’ strain.

Necrosis above the graft union of a 3-yr-old tree of Virginia Crab on MM 104 rootstock infected with the ‘E-36’ strain by double budding (bark removed).
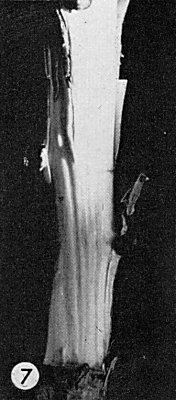
Grooves in the wood of the stem of a 2-yr-old Virginia Crab plant inoculated with the ‘C-431’ strain (bark removed).

Particles of the ‘C-431’ strain stained with uranyl formate. Bar represents 100 nm.
References list for DPV: Apple stem grooving virus (31)
- de Sequeira, Zast. Bilja 16: 247, 1965.
- de Sequeira, Ann. appl. Biol. 60: 59, 1967.
- de Sequeira & Cropley, TagBer. dt. Akad. Landw Wiss. Berl. 97: 35, 1968.
- de Sequeira & Lister, Phytopathology 59: 572, 1969a.
- de Sequeira & Lister, Phytopathology 59: 1740, 1969b.
- de Sequeira & Posnette, In Tech. Commun. Commonw. Bur. Hort. Plantn Crops 30, Suppl. 2/3/4:76a, 1969.
- Lister, Bancroft & Nadakavukaren, Phytopathology 55: 859, 1965.
- Lister, Bancroft & Shay, Phytopathology 57: 819, 1967.
- Mink, Shay, Gilmer & Stouffer, Phytopathology (in press), 1970.
- Waterworth & Gilmer, Phytopathology 59: 334, 1969.