Details of DPV and References
DPV NO: 312 December 1986
Family: Rhabdoviridae
Genus: Cytorhabdovirus
Species: Barley yellow striate mosaic virus | Acronym: BYSMV
Barley yellow striate mosaic virus
R. G. Milne Istituto di Fitovirologia del CNR, Via Vigliani 104, I-10135 Torino, Italy
M. Conti Istituto di Fitovirologia del CNR, Via Vigliani 104, I-10135 Torino, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Conti (1969a) and
Signoret et al. (1972).
Synonyms:
-
Wheat chlorotic streak virus
(Signoret et al., 1976)
- Wheat chlorotic streak mosaic virus (Signoret et al., 1977)
- Cereal striate mosaic virus (Matthews, 1982)
- Wheat chlorotic streak mosaic virus (Signoret et al., 1977)
-
A virus with bacilliform particles c. 55 x 330 nm containing RNA, protein and lipid. Not mechanically transmissible to plants but transmitted in the persistent (propagative) manner by planthoppers. Causes a striate mosaic disease in cereals. Infects plants only in the family Gramineae.
Main Diseases
Causes chlorotic stripes or mosaic in winter wheat in northern Italy (Conti, 1972, 1980) and southern France (Signoret et al., 1972, 1976, 1977). Triticum durum (Fig. 2), T. compactum and T. aestivum are the most susceptible species. The disease is of little economic importance but incidence has ranged from 5 to 8% in winter wheat. Also causes chlorotic striping in maize in Morocco (Lockhart & Elyamani, 1983). Originally transmitted experimentally by naturally infected hoppers to barley but causes no economically important disease in this crop.
Geographical Distribution
Reported from Italy, France, Morocco and Australia, where the MS strain occurs in maize (see Strains).
Host Range and Symptomatology
In Europe, the virus occurs in wheat and Agropyron repens (Conti, 1969a; Signoret et al., 1976) and occasionally in oats and rye (Conti & Milne, 1977; Amici et al., 1978; Conti, 1980). In Morocco, the virus naturally infects maize (Lockhart & Elyamani, 1983). The virus is not transmissible by mechanical inoculation. It was transmitted experimentally by means of the planthopper vector to 26 of 44 species of Gramineae but not to plants in other families (Conti, 1980). Susceptible species were from the genera Lagurus (Agrostideae); Avena and Trisetum (Aveneae); Briza, Bromus, Dactylis and Poa (Festuceae); Hordeum, Lolium, Triticum and Secale (Hordeae); Phalaris (Phalarideae); Sorghum (Andropogoneae); Zea (Maydeae); and Panicum and Setaria (Paniceae). Symptoms (Fig. 1, Fig. 2, Fig. 3) include stunting and mosaic, chlorotic striping, narrowing of leaf blades and apical yellowing, according to the host species or variety (Conti, 1980; Signoret et al., 1972; Lockhart & Elyamani, 1983).
-
Diagnostic species
-
(Symptoms refer to plants hopper-inoculated at the coleoptile stage.)
- Avena sativa (oats) cv. Alba. Striate yellow mosaic, stunting, reddening of basal leaves.
- Bromus aleutensis, B. arvensis and B. rigidus (= B. maximus). Dwarfing and leaf narrowing (Fig. 3).
- Hordeum vulgare (barley) cv. Delisa. Bright yellow striate mosaic, first appearing on third leaf (Fig. 1).
- Setaria italica. Chlorotic leaf stripes, dwarfing.
- Plants reported as not susceptible (Conti, 1980) include Digitaria sanguinalis, Echinochloa crus-galli, Festuca pratensis, Holcus lanatus and Phleum pratense.
- Avena sativa (oats) cv. Alba. Striate yellow mosaic, stunting, reddening of basal leaves.
-
Propagation species
- Hordeum vulgare (barley). Seedlings are inoculated at the coleoptile stage by means of planthoppers, and harvested about 1 month later. Virus-free vectors are maintained on healthy barley and wheat seedlings.
-
Assay species
- No local lesion host is known. Any of the diagnostic species, used at the coleoptile stage, can serve to assay the infectivity of the vector planthopper; barley seedlings are normally used.
Strains
Wheat chlorotic streak virus from France seems indistinguishable from barley yellow striate mosaic virus and is considered the same virus. A Moroccan wheat rhabdovirus is also very similar biologically (Lockhart & Elyamani, 1983) and serologically (Milne et al., 1986) and may be the same. Maize sterile stunt virus from Australia (Greber, 1982, 1983a, 1983b, 1984a, 1984b) is similar to barley yellow striate mosaic virus biologically and reacts with its antiserum (Greber, 1984b) but its nucleocapsid differs serologically by an SDI of about 2 (Milne et al., 1986). The Australian virus should probably be considered a strain (MS) of barley yellow striate mosaic virus intermediate in serological properties between it and northern cereal mosaic virus. Digitaria striate virus resembles barley yellow striate mosaic virus in structure, vector species and cytopathology (Greber, 1979, 1983b) and is related serologically to strain MS although it is biologically distinct (R. S. Greber, personal communication).
Transmission by Vectors
Transmitted propagatively by the planthopper Laodelphax striatellus
(Fig. 4)
(Conti, 1969a,
1969b,
1974,
1980;
Conti & Plumb, 1977;
Leclant & Signoret, 1976;
Signoret et al., 1976,
1977).
A related
planthopper, Javesella pellucida, has transmitted the virus experimentally
though less efficiently
(Conti, 1980).
L. striatellus acquires the virus after
feeding for at least 1 h, becomes infective after a 9-29 day latent period, and
remains infective for life. Hoppers can transmit after inoculation feeding times of
10-15 min or more
(Signoret et al., 1977;
Conti, 1980).
When hoppers are
injected with partially purified virus preparations, about 30% of the survivors become
transmitters after a latent period of 4-6 days
(Conti, 1969b).
In limited tests of transovarial transmission (Conti, 1980), the virus was transmitted by about 9% of the progeny of two of five transmitting L. striatellus females tested. The larvae became infective in their second week and remained so throughout their lives.
Transmission through Seed
No seed transmission detected in wheat cv. Mara (Conti, 1980).
Serology
Moderately immunogenic. Rabbit antisera with titres in gel double diffusion of 1/128 and 1/256 have been prepared (E. Luisoni, unpublished data) by intramuscular injection of partially purified preparations, either treated with a lipid solvent (chloroform) or untreated, respectively. One antiserum of the second kind had a titre of 1/2048 when used for coating homologous nucleocapsids with antibody (Milne et al., 1986). Antisera against the Moroccan isolate (B. F. L. Lockhart, unpublished results) and strain MS (Greber, 1982) have also been prepared.
Relationships
Barley yellow striate mosaic virus,
northern cereal mosaic virus
and
wheat rosette stunt virus
(Anon., 1979;
Zheng et al., 1983;
Kung & Zheng, 1985;
Zheng & Gong, 1985)
are all
rhabdoviruses
of cereals, with similar morphology,
similar or identical planthopper vectors and similar host ranges. Tests for coating of
nucleocapsids with antibody
(Milne et al., 1986)
suggested that northern cereal
mosaic and wheat rosette stunt viruses are closely similar to each other but more
distantly related (SDI=6) to barley yellow striate mosaic virus; they are therefore
considered as two strains or isolates of a virus distinct from barley yellow striate
mosaic virus.
The cytoplasmic rather than nuclear maturation of the virus places it in sub-group 1 of the plant rhabdoviruses.
Stability in Sap
Virus infectivity in barley sap, assayed by injection into L. striatellus, had a thermal inactivation point (10 min) between 50 and 60°C, a dilution end-point between 10-2 and 10-3, and a longevity of 2-4 days at 5°C or 1-2 days at 22°C (Conti, 1980).
Purification
The method of Luisoni (1969), as modified by G. Boccardo and P. Caciagli (unpublished results) is used. Homogenize about 20 g infected barley leaves with 100 ml 0.2 M phosphate buffer, pH 7.2, in the cold, and filter through nylon mesh. Now suspend in the sap extract sufficient Celite to adsorb all or most of the green material; in practice this may be about 10 g, depending on the source of Celite. After stirring for a few minutes, pour the mixture on to a further 50-100 g of Celite previously mixed with buffer and placed in a Buchner funnel of 7.5 cm diameter to form a filter pad. Filter the sap extract through the pad and wash this with a further 100 ml buffer. At least the lowest layer of the pad should remain white. Mix the filtrate with polyethylene glycol (PEG), M. Wt 6000, to 5% (w/v), agitate in the cold for 2 h, and centrifuge at low speed. Resuspend the pellet in 10 ml buffer and, after centrifugation at low speed, float the supernatant fluid on a 20-60% sucrose density gradient formed in the same buffer. After centrifugation (90 min, 100,000 g) recover the light-scattering virus-containing zone, dilute it in the buffer, and ultracentrifuge. Resuspend the pellet in a small volume of 0.02 M phosphate buffer, pH 7.2. After negative staining in uranyl acetate, with or without prior fixation in glutaraldehyde, the product consists of a mixture of undamaged virus particles and others that have a ‘fried egg’ appearance because of partial disruption.
Properties of Particles
Physical properties not determined. When treated with lipid solvents, the envelope is removed to release the nucleocapsids.
Particle Structure
The particles have typical rhabdovirus morphology, being cylindrical with both ends rounded (Fig. 5) and measuring about 55 x 330 nm in uranyl acetate negative stain (Appiano & Conti, 1974; Conti, 1980) and 55 x 270 nm in neutral phosphotungstate (Conti, 1969a). In sections (Fig. 9, Fig. 10) the particle dimensions are 45 x 300-320 nm (Conti & Appiano, 1973; Conti & Plumb, 1977). Unfixed particles partially disrupt in neutral phosphotungstate, and do so also in uranyl acetate when in sap extracts (Fig. 7, Fig. 8) though not when purified. The surface pattern of spikes is not prominent (Fig. 5). The envelope is easily removed with chloroform, 0.2% Nonidet P40 or 0.2% Triton X-100, to leave intact nucleocapsids (Fig. 6). In uranyl acetate, the nucleocapsid measures about 35 x 300 nm and displays obvious cross-striation of pitch 4.4 nm.
Particle Composition
Not determined, but probably very similar to that of
northern cereal mosaic virus
and wheat rosette stunt virus
(Zheng et al., 1983;
Zheng & Gong, 1985;
Gong et al., 1985).
Nucleic acid: RNA
(Bassi et al., 1980).
Protein: No information.
Lipid: Present in the envelope.
Relations with Cells and Tissues
In host plants. Mature virus particles occur in the cytoplasm of parenchyma
and phloem cells, within membrane bound cisternae. ‘Viroplasms’
of fine granular or
fibrous material
(Fig. 9)
form in the cytoplasm, and virus particles appear to bud
from their edges. Virus particles are not found in the nuclei or between the two
nuclear membranes. Spherical vesicles 25-60 nm in diameter are formed within sac-like
structures in the cytoplasm
(Conti & Appiano, 1973).
Enzyme digestion, EDTA
treatment and autoradiography on thin sections of infected barley
(Bassi et al., 1980)
indicate that the virus contains RNA and replicates only in the cytoplasm. The
viroplasms consist mainly of protein. Shortly after inoculation, cytoplasmic RNA
synthesis increases markedly while nuclear RNA synthesis is depressed. After cytoplasmic
RNA synthesis has reached its maximum (12 days post inoculation), viroplasms and virus
particles become visible and macroscopic symptoms appear. The viroplasms are not sites
of increased RNA synthesis, though they may form a reservoir of viral protein
(Bassi et al., 1980).
In host insects. In salivary glands, ovaries and mycetomes of transmitting hoppers, virus particles appear in the cytoplasm but not in nuclei or other organelles. Viroplasms are not seen, but flexuous tubules 20-30 nm wide, enclosed in membranes, occur in the cytoplasm of infected hoppers but not in that of uninfected ones. Their role is not clear. Signs of virus multiplication are not seen in cells of the gut (Conti & Plumb, 1977; Milne et al., 1980).
Notes
Rhabdoviruses
of Gramineae can currently be distinguished by their vector type
(leafhopper or planthopper), particle morphology, site of maturation (nuclear or
cytoplasmic) and serology. Comparative molecular data are scarce. In nucleocapsid
antigenicity and in particle size, the cluster of viruses serologically related to
barley yellow striate mosaic virus differs
(Milne et al., 1986)
from maize mosaic virus
(Falk & Tsai, 1983;
McDaniel et al., 1985)
and from
shiraz maize rhabdovirus
(Izadpanah et al., 1983),
though both,
like barley yellow striate mosaic virus, are transmitted by planthoppers.
Festuca leaf streak virus
is similar in morphology and cytopathology to viruses of the
barley yellow striate mosaic cluster
(van Beek et al., 1985)
though the
vector is unknown; it is apparently not related serologically to barley yellow
striate mosaic virus or
northern cereal mosaic virus
either in the G protein of
the envelope or in the nucleocapsid
(Lundsgaard, 1984).
Two cereal rhabdovirus
diseases known in Siberia,
oat pseudorosette (= zakuklivanie)
and cereal mosaic,
are transmitted by L. striatellus and yield particles that apparently
react with northern cereal mosaic virus antiserum (A. K. Krylov, personal
communication). The oat pseudorosette particles occur in the cytoplasm, associated
with viroplasms; a mycoplasma is also involved in the disease complex
(Fedotina, 1977).
Host range studies indicate some differences between barley yellow striate mosaic
virus and northern cereal mosaic virus. Bromus spp., sweet corn, Poa pratensis,
Sorghum spp. and Trisetum flavescens were found to be susceptible to the
former but not the latter, whereas Alopecurus aequalis and Echinochloa
crus-galli were susceptible to the latter but not the former
(Conti, 1980;
Ito & Fukushi, 1944).
Other rhabdoviruses of Gramineae all have leafhopper vectors (see Conti, 1985; Francki et al., 1985) except cynodon chlorotic streak virus (Lockhart et al., 1985) and possibly finger millet mosaic virus (Maramorosch et al., 1977). However, both these viruses develop within the nucleus or perinuclear space, and are placed in sub-group 2 of the plant rhabdoviruses.
Acknowledgements
Figures 9 and 10 courtesy Dr A. Appiano.
Figures

Infected barley leaf showing characteristic striping.

Infected (above) and healthy (below) leaves of Triticum durum.

Infected (left) and healthy (right) plants of Bromus rigidus.

The planthopper vector, Laodelphax striatellus, female brachypterous form, x 15 (courtesy Dr C. Vidano).

Intact virus particle in uranyl acetate; bar represents 100 nm.

Virus particle in uranyl acetate after treatment with Nonidet P40, showing nucleocapsid only; bar represents 100 nm.

Partly stripped virus particle in uranyl acetate, revealing nucleocapsid at top; bar represents 100 nm.

Damaged virus particle in uranyl acetate, penetrated by stain. Bar represents 100 nm.
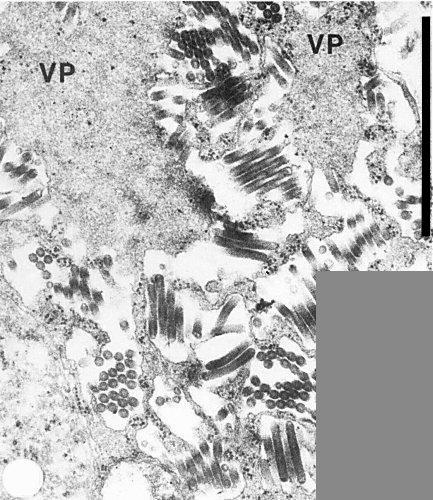
Thin section of infected barley leaf parenchyma showing virus particles associated with viroplasm (VP); the nucleus and nuclear membranes (bottom left) appear unaffected. Bar represents 1 µm.
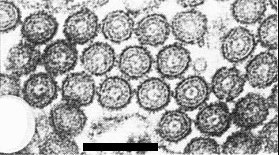
Virus particles in cross section. Bar represents 100 nm.
References list for DPV: Barley yellow striate mosaic virus (312)
- Amici, Faoro & Tornaghi, Riv. Pat. veg., Pavia, Ser. IV 14: 85, 1978.
- Anon., Scientia sin. 22: 573, 1979.
- Appiano & Conti, J. Submicr. Cytol. 6:105, 1974.
- Bassi, Barbieri, Appiano, Conti, D'Agostino & Caciagli, J. Submicr. Cytol. 12: 201, 1980.
- Conti, Phytopath. Z. 66: 275, 1969a.
- Conti, Ricerca scient. 39: 701, 1969b.
- Conti, Phytopath. Z. 73: 39, 1972.
- Conti, Mikrobiologija 11: 49, 1974.
- Conti, Ann. appl. Biol. 95: 83, 1980.
- Conti, in The Leafhoppers and Planthoppers, p. 289, ed. L. Nault & J. Rodrigues, Wiley Interscience, 500 pp., 1985.
- Conti & Appiano, J. gen. Virol. 21: 315, 1973.
- Conti & Milne, Annls. Phytopath. 9: 255, 1977.
- Conti & Plumb, J. gen. Virol. 34: 107, 1977.
- Falk & Tsai, Phytopathology 73: 1536, 1983.
- Fedotina, Arch. Phytopath. PflSchutz 13: 177, 1977.
- Francki, Milne & Hatta, Atlas of Plant Viruses, Vol. 1, CRC Press, Boca Raton, 222 pp., 1985.
- Gong, Zhen, Peng, Tsao & Shikata, Chin. J. Virol. 1: 257, 1985.
- Greber, Aust. J. agric. Res. 30: 43, 1979.
- Greber, Aust. J. agric. Res. 33: 13, 1982.
- Greber, Aust. Microbiol. 4: 100, 1983a.
- Greber, in Proc. int. Maize Virus Dis. Colloq. Workshop, Wooster, Ohio St.Univ., 1982, p. 206, ed. D. Gordon, J. Knoke, L. Nault & R. Ritter, 261 pp.,1983b.
- Greber, Maize Virus Dis. Newsl. 1: 46, 1984a.
- Greber, in Current Topics in Vector Research, Vol. 2, p. 141, ed. K. Harris, Praeger, 1984b.
- Ito & Fukushi, J. Sapporo Soc. Agric. & Forestry 36: 65, 1944.
- Izadpanah, Ahmadi, Parvin & Jafari, Phytopath. Z. 107: 283, 1983.
- Kung (= Gong) & Zheng (= Zhen), Acta Biochim. Biophys. sin. 17: 248, 1985.
- Leclant & Signoret, C. r. IV Journées Phytiatr. Phytopharm. Circum-Méditerr. Montpellier, 1974, p. 215, Rullière-Libeccio, Avignon, 493 pp., 1976.
- Lockhart & Elyamani, in Proc. int. Maize Virus Dis. Colloq. Workshop, Wooster, Ohio St. Univ., 1982, p. 127, ed. D. Gordon, J. Knoke, L. Nault & R. Ritter, 261 pp., 1983.
- Lockhart, Khaless, El Maataoui & Lastra, Phytopathology 75: 1094, 1985.
- Luisoni, Ricerca scient. 39: 708, 1969.
- Lundsgaard, Intervirology 22: 50, 1984.
- Maramorosch, Govindu & Kondo, Pl. Dis. Reptr 61: 1029, 1977.
- Matthews, Intervirology 17: 1, 1982.
- McDaniel, Ammar & Gordon, Phytopathology 75: 1167, 1985.
- Milne, Lenzi & Conti, Abstr. 3rd Conf. Virus Dis. Gramin. Europe, Rothamsted, 1980: 32, 1980.
- Milne, Masenga & Conti, Intervirology 25: 83, 1986.
- Signoret, Giannotti & Alliot, Annls. Phytopath. 4: 45, 1972.
- Signoret, Leclant & Alliot, Poljopr. Znan. Smotra 39(49): 183, 1976.
- Signoret, Conti, Leclant, Alliot & Giannotti, Annls. Phytopath. 9: 381, 1977.
- van Beek, Lohuis, Dijkstra & Peters, J. Ultrastruct. Res. 90: 294, 1985.
- Zheng (= Zhen) & Gong, Chin. J. Virol. 1: 55, 1985.
- Zheng (= Zhen), Xü, Zhu, Shen & Kung (= Gong), Acta Biochim. Biophys. sin. 15: 561, 1983.