Details of DPV and References
DPV NO: 314 December 1986
Family: Potyviridae
Genus: Potyvirus
Species: Yam mosaic virus | Acronym: YMV
Yam mosaic virus
J.-C. Thouvenel Laboratoire de Phytovirologie, Centre ORSTOM d'Adiopodoumé, 01 B.P. V 51, Abidjan, Côte d'Ivoire
C. Fauquet Laboratoire de Phytovirologie, Centre ORSTOM d'Adiopodoumé, 01 B.P. V 51, Abidjan, Côte d'Ivoire
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Thouvenel & Fauquet (1977, 1979).
A virus with flexuous filamentous particles about 785 nm long, consisting of one species of single-stranded RNA and one species of coat protein. It is sap-transmissible with difficulty to a small range of hosts and transmitted by aphids in the non-persistent manner. It causes economically important losses in yams in tropical areas (Africa and America).
Main Diseases
Causes interveinal mosaic and sometimes vein-banding in yams (Dioscorea spp.).
Geographical Distribution
Reported from several African countries: Ivory Coast, Nigeria (Mohamed & Terry, 1979), Togo (Reckhaus, 1980); probably also present in the Caribbean area.
Host Range and Symptomatology
Known to infect several species of Dioscoreaceae, particularly Dioscorea alata, D. cayenensis, D. esculenta and D. rotundata.
-
Diagnostic species
- Dioscorea cayenensis. Systemic symptoms are apt to differ considerably in different
leaves of the same plant: mosaic, vein-banding, green spotting or flecking, curling and
mottling (Fig. 1, Fig. 2,
Fig. 3, Fig. 4,
Fig. 5).
- D. esculenta. Mosaic and vein-banding (Fig. 6).
- D. preussii. Mosaic and vein-banding.
- D. composita and D. floribunda. Not infected.
- Nicotiana benthamiana. Mottling on inoculated and systemically infected leaves.
- N. megalosiphon. Systemic chlorotic spotting on the whole plant.
- D. esculenta. Mosaic and vein-banding (Fig. 6).
-
Propagation species
- N. benthamiana
and D. cayenensis are suitable hosts for maintaining the virus and are good sources of virus for purification.Assay species
- No local lesion assay host is known. D. cayenensis is suitable for assaying transmission by aphids.
Strains
None recorded.
Transmission by Vectors
Transmitted in a non-persistent manner by the aphids Aphis gossypii, A. craccivora, Rhopalosiphum maidis and Toxoptera citricidus. Aphids acquire virus after a 5 min access period and infect plants after an inoculation access period of less than 15 min (Thouvenel & Fauquet, 1979).
Transmission through Seed
No transmission has been detected through seed of Dioscorea cayenensis (Thouvenel & Fauquet, 1979).
Serology
The virus is moderately immunogenic; antiserum with a titre of 1/2048 in microprecipitin tests was obtained. Flocculent precipitates are formed in microprecipitin tests. The virus is detected easily in crude sap of yams by enzyme-linked immunosorbent assay (ELISA) (Thouvenel & Fauquet, 1980).
Relationships
The virus belongs to the potyvirus group. It is serologically related to, but distinguishable from, pepper veinal mottle virus, guinea grass mosaic virus and passionfruit ringspot virus. In Guadeloupe (Caribbean), a virus serologically related to yam mosaic virus and with similar particle shape was reported in Dioscorea trifida (Migliori & Cadilhac, 1976).
Stability in Sap
Crude sap from diseased D. cayenensis leaves was not infective, so the properties of the virus were studied with leaf extracts in phosphate buffer containing cysteine hydrochloride, bentonite and activated charcoal. In such yam extracts, the thermal inactivation point is between 55°C and 60°C; the dilution end-point is 10-2 to 10-3 and longevity in vitro is less than 1 day at 25°C, c. 40 days at 4°C and over 150 days at -30°C.
Purification
Purification of the virus particles from yam leaves is difficult because of the presence of mucilaginous substances; N. benthamiana is a better source of virus for purification. Good results were obtained by the following method: blend fresh infected leaves in 0.2 M potassium phosphate buffer (pH 8) containing 1% mercaptoethanol in the presence of chloroform (2 ml/g leaf). Centrifuge the extract (10 min; 12,000 g) and add ammonium sulphate to 20% (w/v), or PEG, M. Wt 6000, to 6% (w/v) to the supernatant fluid. Centrifuge (15 min; 20,000 g), and resuspend the sediment in water; after a further low speed centrifugation, centrifuge the virus through a ‘cushion’ of 8 ml 20% sucrose in a swinging bucket rotor (2.5 h; 80,000 g). Further purification is obtained by centrifugation in sucrose density gradients. Yield of virus is about 15-25 mg/kg leaf.
Properties of Particles
Purified preparations contain a single sedimenting component.
Sedimentation coefficient (s20, w): c. 150 S (estimated by
centrifugation in sucrose gradients).
Isoelectric point, pHi: 4.3 ± 0.3.
A260/A280: 1.20; Amax :262 nm; Amin
: 247 nm; Amax/Amin : 1.13 (uncorrected for light-scattering).
Particle Structure
The particles are flexuous, with a modal length of 785 + 15 nm and a diameter of 13 nm (Fig. 7).
Particle Composition
Nucleic acid: RNA, single-stranded (determined by polyacrylamide gel electrophoresis after nuclease digestion) comprising c. 6% of the particle weight (based on A260/A280 ratio).
Protein: a single species, of M. Wt c. 34,000 (Thouvenel & Fauquet, 1979). This protein contains about 305 amino acid residues with the following composition: asp 35; thr 15; ser 19; glu 39; pro 14; gly 23; ala 26; cys 3; val 16; met 15; ileu 15; leu 24; tyr 13; phe 11; his 9; lys 16; arg 11; trp 1 (Fauquet & Thouvenel, 1980).
Relations with Cells and Tissues
The virus induces cytoplasmic pinwheel inclusions, scrolls and laminated aggregates which can be detected by electron microscopy (Fig. 8). The virus cannot be eliminated by exposure of infected tubers for 15 min at 55°C.
Notes
Yam mosaic virus differs from dioscorea green-banding virus (Ruppel et al., 1966) in not infecting Crotalaria striata, Nicotiana glutinosa, Dioscorea composita or D. floribunda. A virus from D. trifida in the Caribbean is morphologically similar and serologically related to yam mosaic virus, but there is little other information (Marchoux et al., 1979). Harrison & Roberts (1973) reported a virus of the same size, and not sap-transmissible to plants in families outside the Dioscoreaceae, in D. alata imported into Britain from Barbados; similar particles were found in the leaves of D. esculenta from Sierra Leone. A relationship may exist between these viruses and yam mosaic virus but no serological comparisons have been made.
Figures

Systemic mosaic in leaf of Dioscorea cayenensis.

Systemic dotting in leaf of D. cayenensis.

Systemic flecking in leaf of D. cayenensis.

Systemic vein mosaic in leaf of D. cayenensis.

Systemic vein banding in leaf of D. cayenensis.

Systemic symptom in leaf of D. esculenta.
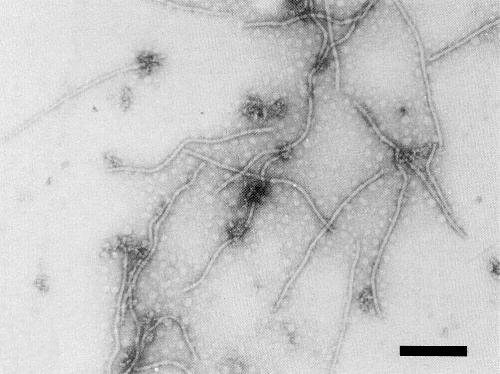
Particles from a purified preparation, negatively stained with 1% uranyl acetate. Bar represents 200 nm.

Electron micrograph of laminated aggregates in section of infected leaf of D. cayenensis. Bar represents 500 nm.
References list for DPV: Yam mosaic virus (314)
- Fauquet & Thouvenel, in Viral Diseases of Crop Plants in Ivory Coast (Init.-Document. Techn. No. 46), p.29, Office de la Recherche Scientifique et Technique Outre-Mer, Paris, 128 pp., 1980.
- Harrison & Roberts, Trop. Agric. Trin. 50: 335, 1973.
- Marchoux, Edwige & Migliori, Annls Phytopath. 11: 59, 1979.
- Migliori & Cadilhac, Annls Phytopath. 8: 73, 1976.
- Mohamed & Terry, Trop. Agric. Trin. 56: 175, 1979.
- Reckhaus, Ph.D. Diss., Univ. Bonn, 103 pp., 1980.
- Ruppel, Delphin & Martin, J. Agric. Univ. P. Rico 50: 151, 1966.
- Thouvenel & Fauquet, C.r. hebd. Séanc. Acad. Sci., Paris 284: 1947, 1977.
- Thouvenel & Fauquet, Ann. appl. Biol. 93: 279, 1979.
- Thouvenel & Fauquet, in L'Igname, p. 101, Les Colloques de l'Institut National de la Recherche Agronomique, Paris, ed. INRA, 296 pp., 1980.