Details of DPV and References
DPV NO: 315 December 1986
Family: Partitiviridae
Genus: Alphacryptovirus
Species: Carnation cryptic virus 1 | Acronym: CCV-1
Carnation cryptic virus
V. Lisa Istituto di Fitovirologia applicata del C.N.R., Via O. Vigliani 104, I-10135 Torino, Italy
E. Luisoni Istituto di Fitovirologia applicata del C.N.R., Via O. Vigliani 104, I-10135 Torino, Italy
R. G. Milne Istituto di Fitovirologia applicata del C.N.R., Via O. Vigliani 104, I-10135 Torino, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Lisa et al. (1980).
A virus with isometric particles about 29 nm in diameter, containing three major size classes of double-stranded (ds) RNA. It occurs symptomlessly in carnation and is present in low concentration in plant tissues. It is apparently not transmissible by sap inoculation or by grafting but is transmitted with high frequency through the seed.
Main Diseases
Causes no symptoms in carnation ( Dianthus caryophyllus) and apparently has no effect on those induced by any other virus when present (Lisa et al., 1981b).
Geographical Distribution
Found in carnation plants from Europe, Israel and USA (Lisa et al., 1981b). Because of the extensive trade in carnation cuttings, the virus is probably present in most carnation growing areas, worldwide.
Host Range and Symptomatology
Found in Mediterranean and miniature or spray carnations (Dianthus caryophyllus) and in commercial hybrids of D. caryophyllus with other Dianthus spp., such as ‘Chinese’ carnations; not found in eleven Sim carnation cultivars. Experimental host range not determined because of lack of transmissibility except through the seed (Lisa et al., 1981b).
- Diagnostic species
No diagnostic species known.
- Assay species
No assay species known.
- Propagation species
Any carrier carnation plant suitable for vegetative propagation, providing that no other carnation virus is present.
Strains
Some isolates from Israel differed serologically from the original virus described in Italy (see Relationships).
Transmission by Vectors
No vector known. An attempt to transmit the virus from carnation to carnation by Myzus persicae was inconclusive (Lisa et al., 1980).
Transmission through Seed
Transmitted with high frequency through seeds of Mediterranean and hybrid carnations: the virus was detected in 90 to 100% of seedlings of Mediterranean and hybrid ‘Chinese’ carnation cultivars. In the garden carnation cv. ‘Chabaud’ the virus was found in c. 10% of the seedlings checked. All the seedlings tested (about 200) were from parents whose virus status was unknown (Lisa et al., 1981b).
Transmission by Grafting
Not transmitted from carnation to carnation either by grafting carrier scions to non-carrier stocks or vice versa, even when the grafts took well and survived up to nine months (Lisa et al., 1981b). Healthy carnation plants grafted with scions carrying carnation cryptic virus and also carnation mottle and carnation etched ring viruses had not acquired carnation cryptic virus after 6 months although the two other viruses crossed the graft union (V. Lisa, E. Luisoni & R. G. Milne, unpublished data).
Serology
The virus is a relatively good immunogen and antisera with titres of 1/512 in agar gel double diffusion tests have been obtained. Owing to the low virus content in carrier plants, concentrated sap is required to obtain reliable results with this test. Double antibody sandwich ELISA and immunoelectron microscopy (immunosorbent electron microscopy (ISEM) plus decoration) detect the virus in crude plant sap (Lisa et al., 1981b).
Relationships
Some isolates from Israel differed serologically from the original virus described in Italy. In tests with antiserum to the original isolate, the isolates from Israel gave very low ELISA values and low particle counts in ISEM (only 20% of those given by the homologous virus). In decoration tests, the titre of the antiserum to the heterologous particles was two twofold dilution steps lower than that to the homologous particles (J. Cohen, M. Alper, R. G. Milne & P. Roggero, unpublished data, cited in Boccardo et al., 1987). The virus has many properties in common with other cryptic viruses (Francki et al., 1985; Boccardo et al., 1987; Descr. No. 298). However it was not serologically related to the following cryptic viruses: beet cryptic 1 and 2, poinsettia cryptic, ryegrass cryptic, white clover cryptic 1 and 2, and the three cryptic viruses present in hop trefoil (Boccardo et al., 1987). Cryptic viruses also share some properties with certain mycoviruses, especially those of subgroup D (Buck et al., 1984). No reaction was found between carnation cryptic virus and 10 antisera to 15 mycoviruses (E. Luisoni and R. G. Milne, unpublished data; antisera listed in Boccardo et al., 1987). There was also no serological relationship to any of 25 isometric viruses with single-stranded (ss) RNA genomes (Lisa et al., 1981b).
Stability in Sap
Thermal inactivation, longevity in vitro and dilution end-point have not been determined because infectivity tests are not feasible. The virus is rather stable in vitro in its serological and morphological properties (Lisa et al., 1981b).
Purification
The virus particles can survive different purification procedures, but the yield may be only 50-100 µg per kg fresh plant tissue, because of the low concentration of virus in the plants. A satisfactory purification procedure is as follows: homogenize 500 g carnation leaves and shoots in 1.5 litres 0.5 M potassium phosphate buffer, pH 7, containing 0.01 M sodium diethyldithiocarbamate, 0.02 M sodium sulphite and 0.005 M disodium ethylenediamine-tetraacetate, emulsify the extract with an equal volume of chloroform, separate the phases by low speed centrifugation and pellet the virus from the aqueous phase by ultracentrifugation. Resuspend the sediments in 0.5 M potassium phosphate buffer, pH 7, containing 0.02 M sodium sulphite and repeat the chloroform treatment and the high speed centrifugation. Resuspend the sediments in 0.05 M phosphate buffer, pH 7, and centrifuge at low speed. To recover as much virus as possible, resuspend and reprecipitate the low speed pellet several times, pooling the supernatant fluids, to obtain a volume up to 1/100 that of the initial extract. Ultracentrifuge the preparation for 2-4 h through a preformed 20-50% caesium sulphate density gradient prepared in the same buffer as above. Collect the virus band or bands (Fig. 1) and remove the caesium sulphate by dialysis or ultracentrifugation (Lisa et al., 1981b and unpublished data).
Properties of Particles
After isopycnic centrifugation in caesium chloride in 0.01 M phosphate buffer, pH 7, the virus formed a major band with buoyant density 1.337 g/ml plus other minor bands that varied in position and density in different preparations (Lisa et al., 1981b).
Particle Structure
Purified virus particles negatively stained in uranyl acetate appear rounded in profile, about 29 nm in diameter, and partly penetrated by the stain (Fig 2, Fig. 3). No clear subunit structure is visible. The particles appear similar in other negative stains such as neutral phosphotungstate or ammonium molybdate, pH 7 (Lisa et al., 1981b).
Particle Composition
Nucleic acid. Three major size classes of dsRNA of M. Wt (x 106) 1.04, 0.95 and 0.84 (Fig. 6), plus a minor component of M. Wt 0.88 x 106, as determined by electrophoresis in 5% polyacrylamide gels in non-denaturing conditions (Lisa et al., 1981a). The minor component was not detected in all preparations. Purified RNA preparations reacted with a serum containing antibodies to dsRNA (Fig. 5). In the electron microscope the dsRNA molecules appear linear under non-denaturing conditions, whereas the ssRNA molecules of carnation mottle virus have obvious secondary structure (Fig. 7, Fig. 8) (Lisa et al., 1981a). RNA preparations from carnation cryptic virus did not hybridize in Northern blot experiments with cDNA to RNA preparations extracted from white clover cryptic viruses 1, 2 or 3, or beet cryptic virus 1 or 2 (G. Boccardo and P. Palukaitis, personal communication).
Protein: No information available.
Relations with Cells and Tissues
In thin sections of carrier carnation plants, virus particles were not seen, possibly because of their low concentration and similarity to ribosomes. No cytopathological disorders were associated with the presence of the virus (Lisa et al., 1981b).
Notes
Some clones of carnation contain a second dsRNA virus of similar size but apparently serologically unrelated to carnation cryptic virus (V. Lisa, R. G. Milne and G. Boccardo, unpublished data). Mediterranean carnations may be infected by the ‘attenuated form’ of carnation mottle virus. This occurs in low concentration, like a cryptic virus, but may be distinguished by infectivity and serological tests on concentrated preparations (Fig. 4) (Luisoni et al., 1984). Antisera to other carnation viruses, if prepared using carnation as virus source, may contain antibodies to carnation cryptic virus and so give rise to spurious results; the presence of dsRNA in the particles may elicit antisera cross-reacting with other dsRNA viruses. These possibilities should be considered when doing serological tests. The probably wide distribution of carnation cryptic virus in carnations may also complicate screening for other viruses based on detection of dsRNA in the plants.
Carnation cryptic virus was not eliminated from four Mediterranean carnation cultivars by heat therapy (37°C for 4 weeks), followed by meristem tip culture (A. Baratta, E. Luisoni & V. Lisa, unpublished data).
Carnation cryptic virus particles contain three major RNA size classes whereas several other cryptic viruses contain only two. It is not yet determined whether all three RNA species are genomic or whether one or more are satellites or subgenomic sequences.
Morphologically, cryptic viruses can be divided into two types, those with rather featureless particles 29-32 nm in diameter, and those with particles about 38 nm in diameter possessing prominent subunits (Boccardo et al., 1987). Carnation cryptic virus belongs to the first type.
Figures

Light-scattering band (arrow) formed by carnation cryptic virus after 4 h centrifugation at 149,800 g in a 20-50% preformed caesium sulphate density gradient.

Purified particles of the virus in uranyl acetate. Bar represents 100 nm.
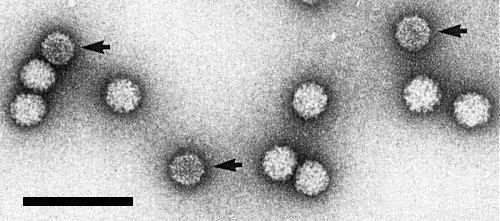
Particles of carnation cryptic virus (arrows) mixed with those of tomato bushy stunt virus (diameter 34 nm). Bar represents 100 nm.

Concentrated preparation from carnations containing particles of carnation cryptic virus (left) and the attenuated form of carnation mottle virus (right) decorated with carnation mottle virus antibody. Bar represents 100 nm.
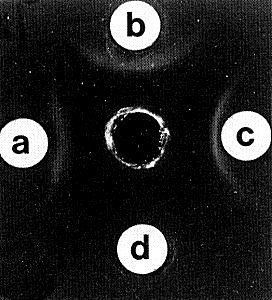
Reaction of a maize rough dwarf virus antiserum containing antibodies to dsRNA (Luisoni et al., 1975) with: (a) synthetic poly(I): poly(C), (b) carnation cryptic virus dsRNA, (c) rice gall dwarf virus dsRNA; (d) shows non-reaction with tobacco mosaic virus ssRNA.
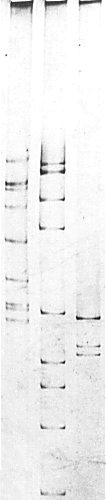
Electrophoretic separation in a 5% polyacrylamide gel slab of dsRNA species from maize rough dwarf virus (left), Bombyx mori cytoplasmic polyhedrosis virus (middle), and carnation cryptic virus (right).

Nucleic acid from purified particles of carnation cryptic virus spread under non-denaturing conditions and shadowed. Note the extended linear molecules differing from those in Fig. 8. Bar represents 100 nm.

Preparation as in Fig. 7 of RNA from carnation mottle virus. Note that the molecules are not extended but have a ‘collapsed’ appearance indicating extensive secondary structure. Bar represents 100 nm.
References list for DPV: Carnation cryptic virus (315)
- Boccardo, Milne, Luisoni, Lisa & Accotto, Virology 147: 29, 1985.
- Boccardo, Lisa, Luisoni & Milne, Adv. Virus Res. 32: 171, 1987.
- Buck, Ackermann, Bozarth, Bruenn, Koltin, Rawlinson, Ushiyama & Wood, Intervirology 22: 17, 1984.
- Francki, Milne & Hatta, Atlas of Plant Viruses, Vol. II, 284pp., CRC Press, Boca Raton, Florida, 1985.
- Lisa, Luisoni & Milne, Acta Hort. 110: 175, 1980.
- Lisa, Boccardo & Milne, Virology 115: 410, 1981a.
- Lisa, Luisoni & Milne, Ann. appl. Biol. 98: 431, 1981b.
- Luisoni, Milne & Boccardo, Virology 68: 86, 1975.
- Luisoni, Lisa & Roggero, Atti Giornate Fitopatol. 1984 3: 427, 1984.