Details of DPV and References
DPV NO: 318 December 1986
Family: Unallocated ssRNA+ viruses
Genus: Sobemovirus
Species: Solanum nodiflorum mottle virus | Acronym: SNMoV
Solanum nodiflorum mottle virus
R. S. Greber Agricultural Research Laboratories, Department of Primary Industries, Indooroopilly 4068, Australia
J. W. Randles Waite Agricultural Research Institute, The University of Adelaide, South Australia 5064, Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Greber (1965;
1973).
Selected synonyms
- Solanum nigrum mosaic virus
(Greber, 1965)
- Solanum nodiflorum virus (Hollings et al., 1979)
-
A virus with isometric particles 28-30 nm in diameter occurring commonly in Solanum nodiflorum in eastern Australia. The virus has a narrow host range, but is stable and readily transmitted by inoculation of sap. In natural infections the virus particles contain a linear genomic RNA and a circular single-stranded satellite RNA. The virus is transmitted in nature by coccinellid beetles.
Main Diseases
Naturally occurring isolates of the virus containing satellite RNA cause a prominent light and dark green mottle with associated rugosity in leaves of Solanum nodiflorum (= S. americanum; Henderson, 1974; Purdie et al., 1982). The virus occurs less commonly in S. nigrum and S. nitidibaccatum (= S. sarrachoides; Purdie et al., 1982) but causes similar symptoms. Incidence increases through the summer when vectors are active.
Geographical Distribution
The virus appears to be endemic to Australia. It has been reported only from eastern Australia between latitudes 16 and 30°S and within 300 km of the coast, generally coinciding with the distribution of its main host and its beetle vectors. The geographical distributions of the virus and its wild plant host do not overlap with those of the serologically related but distinct velvet tobacco mottle virus.
Host Range and Symptomatology
The natural host range is restricted to three Solanum species, of which at least one (S. nodiflorum) is probably native to Australia (Henderson, 1974) and all are good hosts of the three species of coccinellid known to be vectors. The experimental host range of the virus is narrow and the virus may be difficult to recover from some species. Systemic infection occurs in S. nodiflorum, S. nitidibaccatum, S. nigrum and S. villosum following inoculation by sap or vector. Sap inoculation produces systemic infection of Nicotiana benthamiana, N. clevelandii and Tetragonia expansa. Additional hosts from which virus has been recovered from inoculated leaves are Chenopodium quinoa, Datura stramonium, Gomphrena globosa, Nicandra physalodes, Nicotiana debneyi, N. glutinosa, N. tabacum, N. velutina, Solanum melongena and S. rostratum (Greber, 1981; Jones & Mayo, 1984).
-
Diagnostic species
- Solanum nodiflorum.
Vein clearing of young leaves appears after c. 15 days but a severe epinasty of these leaves occurs when the plants are inoculated by the vector. Systemically infected leaves develop a prominent light and dark green mottle with associated puckering (Fig. 1). - Nicotiana clevelandii. A few diffuse chlorotic lesions may show on
inoculated leaves; new
growth shows vein chlorosis, then puckering and systemic mottle
(Fig. 2).
- N. debneyi. Necrotic local lesions and sometimes small chlorotic ring lesions develop under certain conditions (Fig. 3, Fig. 4) (Jones & Mayo, 1984).
- Solanum nodiflorum mottle virus and velvet tobacco mottle virus can be differentiated by exclusive systemic infection of their major natural hosts, S. nodiflorum or N. velutina, respectively.
- N. debneyi. Necrotic local lesions and sometimes small chlorotic ring lesions develop under certain conditions (Fig. 3, Fig. 4) (Jones & Mayo, 1984).
-
Propagation species
- N. clevelandii
is the best source of virus for purification, but S. nodiflorum is useful for virus culture and vector maintenance.Assay species
- N. debneyi
is the only useful local lesion host (Jones & Mayo, 1984). Non-quantitative infectivity tests can be done on S. nodiflorum.
Strains
No serological or host range differences have been found among natural isolates. Local lesion isolates have been obtained that do not contain the satellite RNA (Jones & Mayo, 1984).
Transmission by Vectors
Following the terminology of Richards (1983), the established vectors (Greber, 1981) are Epilachna vigintioctopunctata pardalis (= E. sparsa) (Fig. 5), E. vigintisexpunctata vigintisexpunctata (= E. doryca australica) and E. guttatopustulata. All three vector species transmit the virus efficiently during the first feed after a 24 h acquisition access period, but with a 2 h acquisition period efficiency drops to c. 50% for single insects. Insects remain able to transmit for only 10 h if transferred to new plants at 2 h intervals, but for more than 24 h when fasting. Larvae are as efficient vectors as adults (Greber, 1981) but virus acquired by larvae is not transmitted when they become adults. A Psylliodes sp. (flea beetle) is an inefficient vector. The mirid vector of velvet tobacco mottle virus, Cyrtopeltis nicotianae, also transmits solanum nodiflorum mottle virus (J. Randles, unpublished data), but does not seem to be an important natural vector (Greber, 1981).
Transmission through Seed
Not found for S. nodiflorum or N. clevelandii (R. Greber, unpublished data).
Serology
The virus is an efficient immunogen, and rabbit antisera with titres of 1/1000-1/2000 in gel diffusion tests, and giving a single precipitation line, are readily prepared (Hollings et al., 1979; Greber, 1981).
Relationships
The virus is serologically related to velvet tobacco mottle virus, high-titred antisera giving an SDI of c. 2 (Randles et al., 1981; Chu & Francki, 1983), and substantial spurs being produced in gel diffusion comparisons (Fig. 14). No serological relationship was found to 53 other viruses with isometric particles (Hollings et al., 1979). Despite the serological relationship between solanum nodiflorum mottle virus and velvet tobacco mottle virus, the particle coat proteins have different M. Wt (solanum nodiflorum mottle virus, 31,000; velvet tobacco mottle virus, 37,000). Nevertheless these proteins may originate from a similarly sized precursor because solanum nodiflorum mottle virus RNA specifies an in vitro translation product of M. Wt 38,000 (Kiberstis & Zimmern, 1984) and the 37,000 M. Wt protein of velvet tobacco mottle virus is unstable, degrading to a 32,000 M. Wt protein (Chu & Francki, 1983). Analysis by cDNA hybridization indicates that sequence homology for the RNA1 molecules of these viruses is 20%-50%, depending on the stringency of the conditions (Gould & Hatta, 1981), whereas the sequence homology for their satellite RNA species is 92-95% (Haseloff & Symons, 1982).
Solanum nodiflorum mottle virus resembles sobemoviruses (Tremaine & Hamilton, 1983) in particle morphology, coat protein M. Wt, size and type of genome RNA, ease of mechanical transmission, high thermal stability in vitro, narrow host range, particle density and sedimentation rate, presence of particles in both cytoplasm and nucleus, and beetle transmission.
Two other viruses that occur in Australia, lucerne transient streak virus and subterranean clover mottle virus, also have sobemovirus characteristics and circular satellite RNA species. The satellite RNA species of solanum nodiflorum mottle virus and velvet tobacco mottle virus can multiply in association with lucerne transient streak virus (Jones & Mayo, 1983; 1985) and the satellite RNA of lucerne transient streak virus can multiply in association with southern bean mosaic virus (Paliwal, 1984). This provides further evidence that solanum nodiflorum mottle virus, velvet tobacco mottle virus and lucerne transient streak virus are sobemoviruses.
Stability in Sap
The virus is infective in N. clevelandii sap after dilution in excess of 10-5, after heating for 10 min at 90°C, and on storage for 70 days at 24°C, 1 year at 2°C or 7 years at -20°C (Greber, 1981).
Purification
N. clevelandii plants, harvested 3-4 weeks after inoculation, are the best virus source and clarification of tissue extracts by n-butanol (8.5-9%, v/v) or a 1:1 mixture of chloroform and n-butanol is effective (Greber, 1981; Hollings et al., 1979; Kiberstis & Zimmern, 1984). Tissue can be stored frozen before extraction in 0.1 M phosphate buffer, pH 7.0, with 3 mM EDTA and antioxidant. A polyethylene glycol (PEG), M. Wt 6000, precipitation step can be included (8-10% PEG + 1-3% NaCl) (Greber, 1981; Kiberstis & Zimmern, 1984). The virus particles are then banded by rate zonal centrifugation in a 10-40% sucrose gradient or by isopycnic centrifugation in caesium chloride solution with a starting density of 1.35 g/cm3. Yields of 0.2 mg/g tissue are obtained.
Properties of Particles
The virus particles sediment as a single component.
Sedimentation coefficient, s20,w: 114 S (Hollings et al., 1979).
A260/A280: 1.53; Amax/Amin: 1.31 (Greber, 1981).
Absorption coefficient, A260(0.1%, 1cm): c. 5.0.
Electrophoretic mobility in 0.8% agar gel of pH 7.6: -22.3 x 10-6 cm sec-1 volt-1 (Hollings et al., 1979).
The infectivity of a 500 µg/ml suspension persisted after treatment with diethyl ether (20%, 24 h), precipitation with ethanol (50% v/v) or ammonium sulphate (30% w/v) (Greber, 1981) but was inactivated by sonication (100 W, 5 min) or by treatment with sodium orthophosphate (2%, 5 min).
Particle Structure
The particles are isometric, 28 nm between opposite faces when measured in compact hexagonal arrays negatively stained with 1% potassium phosphotungstate (Fig. 6). In thin sections the spacing of particles in crystalline arrays is c. 25 nm (Fig. 8). Tubular structures of the same diameter are often associated with the isometric particles (Greber, 1981) (Fig. 7, Fig. 9, Fig. 10).
Particle Composition
Nucleic acid: The particles contain an abundant ssRNA (RNA1) of M. Wt c. 1.5 x 106 (c. 4.5 kb). In addition, particles of all naturally occurring isolates studied contain a satellite RNA species of M. Wt c. 0.13 x 106, which exists in both circular (RNA2) and linear (RNA3) forms (Gould & Hatta, 1981; Jones & Mayo, 1984; Fig. 12, Fig. 13). Isolates that lack satellite RNA have been obtained by single lesion isolation techniques (Jones & Mayo, 1984). The satellite RNA contains 378 nucleotides (Haseloff & Symons, 1982; Keese & Symons, 1987; Fig. 15), forming a covalently closed loop which is partially base-paired; it melts co-operatively with a Tm of 38°C in 10 mM NaCl, 1 mM Na cacodylate, 0.1 mM EDTA, pH 6.8 (Randles et al., 1982). A 2' phosphomonoester group with the core structure C(2'p)(3'p5'A) occurs at residue number 49 in the sequence and is the probable ligation site for circularization of the molecule (Kiberstis et al., 1985).
Protein: The coat protein M. Wt is 30,000-31,000 (Hollings et al., 1979; Greber, 1981; Chu & Francki, 1983). A minor component of M. Wt 28,000 has been reported. Virus RNA (RNA1) specifies an in vitro translation product of M. Wt 38,000 (Kiberstis & Zimmern, 1984), which is recognized by antiserum to the 30,000 M. Wt particle protein. The proteins of purified particles are antigenically similar to those of velvet tobacco mottle virus but chemically distinct (Chu & Francki, 1983).
Genome Properties
The production experimentally of virus isolates that lack RNA2 and RNA3 indicates that in these isolates RNA1 is infective alone and that RNA2 is a satellite (Jones & Mayo, 1984). This leaves unexplained the results of Gould et al. (1981), who found that RNA1 was non-infective and dependent on RNA2 for infectivity. RNA2 is not known to be infective alone (Gould et al., 1981).
Relations with Cells and Tissues
Virus particles occur in the cells of most tissues (Greber, 1981). They are often scattered throughout the cytoplasm and nucleus, and sometimes form crystalline arrays or are constrained into compact groups by membranes (Fig. 7, Fig. 8, Fig. 9, Fig. 10). The largest cytoplasmic accumulations were found in cells with large nuclei and small or no vacuoles. Long tubular or rod-shaped structures were sometimes associated with these particles, whereas shorter rod-shaped structures were mixed with the isometric particles in some nuclei (Fig. 10).
Notes
The major natural hosts of solanum nodiflorum mottle virus and velvet tobacco mottle virus are exclusive, i.e. solanum nodiflorum mottle virus does not systemically infect N. velutina and velvet tobacco mottle virus does not systemically infect S. nodiflorum. In addition, there is no overlap of the natural geographical range of the viruses, and the range of the beetle vectors of solanum nodiflorum mottle virus does not encroach on that of velvet tobacco mottle virus (R. S. Greber, unpublished data). The rods associated with the virus particles in thin sections (Greber, 1981) resemble those associated with rice yellow mottle virus (Bakker, 1975).
Figures

Mottle symptoms in Solanum nodiflorum.

Systemically infected leaves of Nicotiana clevelandii.

Local lesions in Nicotiana debneyi leaves: heated glasshouse, 18 h day. (From Jones & Mayo, 1984.)

Local lesions in Nicotiana debneyi leaves: 5 days at 25°C, then as for Fig. 3. (From Jones & Mayo, 1984.)

Larvae (above) and adults (below) of Epilachna vigintioctopunctata pardalis.

Particles negatively stained with potassium phosphotungstate. Bar represents 70 nm.

Arrays of particles and rod structures in the cytoplasm of a cell of infected S. nodiflorum. Bar represents 200 nm.
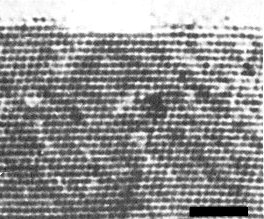
Paracrystalline array of particles in cytoplasm. Bar represents 200 nm.

Rod-shaped or tubular structures and particles in cytoplasm. Bar represents 200 nm.
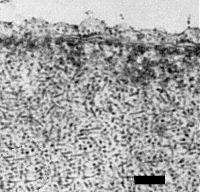
Particles and short rods in nucleus of infected N. clevelandii cell. Bar represents 200 nm.

Electron micrograph of purified RNA1. Bar represents 200 nm. (Courtesy A. R. Gould.)

Electron micrograph of circular satellite RNA. Bar represents 200 nm. (Courtesy A. R. Gould.)

Polyacrylamide gel separation of RNA1 (1) and circular (2C) and linear (2L) forms of RNA2 in a 3.3% polyacrylamide, 8 M urea gel.

Gel-diffusion serology with solanum nodiflorum mottle virus antiserum (A). Note formation of spurs between solanum nodiflorum mottle virus (S) and velvet tobacco mottle virus (V). Virus-free sap (H).
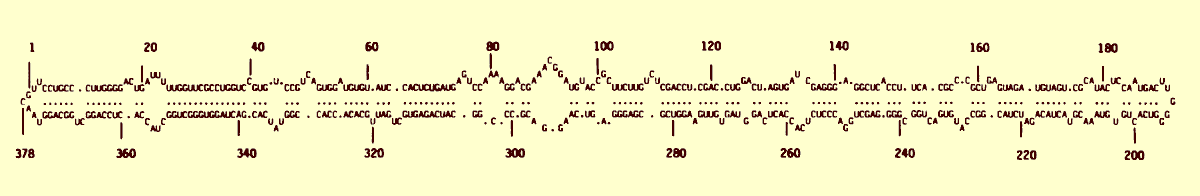
Nucleotide sequence diagram of solanum nodiflorum mottle virus satellite RNA (J02386) with predicted secondary structure (redrawn from Haseloff & Symons, 1982).
References list for DPV: Solanum nodiflorum mottle virus (318)
- Bakker, CMI/AAB Descr. Pl. Viruses 149, 4 pp., 1975.
- Chu & Francki, Virology 129: 350, 1983.
- Gould & Hatta, Virology 109: 137, 1981.
- Gould, Francki & Randles, Virology 109: 420, 1981.
- Greber, Aust. Pl. Dis. Rec. 17: 18, 1965.
- Greber, Aust. Pl. Path. Soc. Newsl. 2: 3, 1973.
- Greber, Aust. J. biol. Sci. 34: 369, 1981.
- Haseloff & Symons, Nucleic Acids Res. 10: 3681, 1982.
- Henderson, Qd Herb. Contrib. 16: 78 pp., 1974.
- Hollings, Stone, Barton & Greber, Rep. Glasshouse Crops Res. Inst., 1978: 150, 1979.
- Jones & Mayo, J. gen. Virol. 64: 1771, 1983.
- Jones & Mayo, J. gen. Virol. 65: 1713, 1984.
- Jones & Mayo, Rep. Scott. Crop Res. Inst. 1984: 190, 1985.
- Keese & Symons, in Viroids and Viroid-like Pathogens, ed. J. Semancik, Boca Raton: CRC Press, 1987.
- Kiberstis & Zimmern, Nucleic Acids Res. 12: 933, 1984.
- Kiberstis, Haseloff & Zimmern, EMBO Jl 4: 817, 1985.
- Paliwal, Can, J. Pl. Path. 6: 93, 1984.
- Purdie, Symon & Haegi, Flora of Australia Vol. 29, 208 pp., 1982.
- Randles, Davies, Hatta, Gould & Francki, Virology 108: 111, 1981.
- Randles, Steger & Riesner, Nucleic Acids Res. 10: 5569, 1982.
- Richards, Int. J. Ent. 25: 11, 1983.
- Tremaine & Hamilton, CMI/AAB Descr. Pl. Viruses 274, 6 pp., 1983.