Details of DPV and References
DPV NO: 321 December 1986
Family: Tombusviridae
Genus: Dianthovirus
Species: Sweet clover necrotic mosaic virus | Acronym: SCNMV
Sweet clover necrotic mosaic virus
C. Hiruki Department of Plant Science, University of Alberta, Edmonton, Alberta, T6G 2P5, Canada
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Hiruki et al. (1984c).
A virus with isometric (icosahedral) particles 34 nm in diameter which contain one or other of two species of single-stranded RNA. Both RNA species are essential for infectivity. The virus is transmitted by contact and by drainage water.
Main Diseases
Causes mosaic, ringspots and systemic veinal necrosis (Fig. 1) in leaves of sweet clovers (Melilotus officinalis and M. alba), resulting in stunting and distortion (Hiruki et al., 1984c; Hiruki, 1986). Also found in alfalfa (Inouye & Hiruki, 1985).
Geographical Distribution
Reported from Alberta, Canada (Hiruki et al., 1981, 1984c) in sweet clovers growing in regions of dark grey soil and black soil (Hiruki, 1986).
Host Range and Symptomatology
Sixteen of 25 species in six dicotyledonous families were infected by inoculation with sap (Hiruki et al., 1984c).
-
Diagnostic species
- Chenopodium amaranticolor.
Small necrotic local lesions in 2-3 days (Fig. 2). No systemic infection. - C. quinoa. Necrotic local lesions in 2-3 days. No systemic infection.
- Cucumis sativus (cucumber) cv. National Pickling. Necrotic local lesions in cotyledons. No systemic infection.
- Gomphrena globosa. Necrotic local lesions in 2-3 days. No systemic infection.
- Nicotiana clevelandii. Necrotic local lesions and necrotic rings in 3-6 days. Systemic chlorotic rings, mottle, leaf distortion and stunting.
- N. glutinosa. Symptomless infection of inoculated leaves. Systemic mild mosaic.
- N. tabacum (tobacco). A few necrotic local lesions in cv. Havana 425. Symptomless infection of inoculated leaves in cv. Bright Yellow. No systemic infection.
- Phaseolus vulgaris (French bean) cv. Red Kidney. Numerous necrotic local lesions, often forming ringspots, in 2-3 days (Fig. 4), enlarging and coalescing to form necrotic areas. Systemic but irregular necrosis, resulting in leaf distortion. Stunting often follows.
- Vicia faba (broad bean) cv. Broad Windsor. Necrotic ringspots in inoculated leaves. No systemic infection.
- Vigna unguiculata (cowpea) cv. Early Ramshorn. Reddish-brown local lesions (Fig. 3), often followed by systemic mottle and apical necrosis.
- Cucumis sativus (cucumber) cv. National Pickling. Necrotic local lesions in cotyledons. No systemic infection.
-
Propagation species
- Nicotiana clevelandii
and Phaseolus vulgaris are useful for virus propagation.Assay species
- Chenopodium amaranticolor
and C. quinoa are good local lesion hosts.
Strains
Isolates from sweet clover and alfalfa can be distinguished by spur formation in gel diffusion tests and by differences in host reaction (Inouye & Hiruki, 1985).
Transmission by Vectors
The virus is exuded into soil from roots of infected plants and can infect adjacent healthy plants. Olpidium brassicae does not serve as a vector (Hiruki, 1986).
Transmission through Seed
None found in seed of infected Vigna unguiculata cv. Early Ramshorn.
Serology
The virus is a good immunogen and antigen in vitro. Antisera giving ring interface test titres of 1/1024 and gel diffusion precipitin titres of up to 1/512 have been obtained. Virus can be detected by both double antibody sandwich and plate-trapped antigen forms of ELISA (Hiruki et al., 1984b; Hiruki, 1986). Immunosorbent electron microscopy was useful in determining the serological specificity of dianthoviruses and their genome pseudo-recombinants (Chen et al., 1984). Monoclonal antibodies specific to the virus were produced (Hiruki, 1982; Hiruki et al., 1984a). Using these antibodies in indirect ELISA (initial plate coating with antigen: antiviral monoclonal antibody; goat anti-mouse IgG coupled with alkaline phosphatase), carnation ringspot virus, sweet clover necrotic mosaic virus and several strains of red clover necrotic mosaic virus have been differentiated (Hiruki & Figueiredo, 1985).
Relationships
Sweet clover necrotic mosaic virus is similar to carnation ringspot virus and red clover necrotic mosaic virus in physical and chemical properties and these three viruses form the dianthovirus group (Matthews, 1982; Gould et al., 1981; Hiruki et al., 1984c); only distantserological relationships were found between them (Hiruki et al., 1984b, 1984c; Hiruki & Figueiredo, 1985). Each of the viruses can form stable genome pseudo-recombinants with the others (Hiruki et al., 1981; Okuno et al., 1983; Lommel, 1983).
Their ability to form pseudo-recombinants indicates that sweet clover necrotic mosaic virus, red clover necrotic mosaic virus and carnation ringspot virus share a common gene pool and might suggest that these viruses are strains of one virus. However, sweet clover necrotic mosaic virus shows only a distant serological relationship to carnation ringspot virus in the plate-trapped antigen form of ELISA (Hiruki et al., 1984b). Moreover, there is no cross-reactivity between sweet clover necrotic mosaic virus and carnation ringspot virus in agar gel double diffusion tests, ring interface tests (Hiruki et al., 1984c), or immunosorbent electron microscopy (Chen et al., 1984). Sweet clover necrotic mosaic virus and carnation ringspot virus can also be distinguished by symptomatology in several indicator hosts such as P. vulgaris cv. Red Kidney, C. amaranticolor, C. sativus and G. globosa (Rao & Hiruki, 1985b). Serological relationships between sweet clover necrotic mosaic virus and strains of red clover necrotic mosaic virus too are remote, their serological differentiation indexes being 7 in ring interface tests (Hiruki et al., 1984c), 5 in agar gel double diffusion tests (Hiruki et al., 1984c), 7 to 10 in passive haemagglutination inhibition tests and 9 in reverse passive haemagglutination inhibition tests (Hiruki et al., 1984b).
Stability in Sap
In sap of Phaseolus vulgaris the thermal inactivation point (10 min) is about 90°C and the dilution end-point between 10-6 and 10-7. Infectivity survived in air-dried leaf tissue for 10 weeks at 22°C (Hiruki, 1986).
Purification
The virus is readily purified from systemically infected Nicotiana clevelandii or from inoculated leaves of Phaseolus vulgaris (Hiruki et al., 1984c): harvest plants 2-3 weeks (N. clevelandii) or 5-6 days (P. vulgaris) after inoculation, homogenize frozen leaves in cold 0.1 M phosphate (Na2HPO4-KH2PO4) buffer, pH 7.0 (1:2, w/v) containing 0.5% ascorbic acid, and filter through cheesecloth. Clarify the sap by adding 1 vol of a 1:1 mixture of chloroform and n-butanol to 1 vol. of sap and then precipitate the virus by adding polyethylene glycol, M. Wt 6000, to 8% (w/v) and sodium chloride to 0.4% (w/v), followed by two cycles of differential centrifugation (108,000 g for 120 min, 9000 g for 10 min). Purify further by sucrose density gradient centrifugation. Yields up to 300 mg/kg leaf tissue can be obtained.
Properties of Particles
Purified preparations usually produce a single light-scattering band in density-gradient centrifugation. In analytical ultracentrifugation, a single component is obtained.
Sedimentation coefficient, s°20, w = 126 S (Hiruki et al., 1984c).
A260/A280: 1.80 (value uncorrected for light-scattering).
Particle Structure
In 2% phosphotungstate (pH 7.0) the virus particles are isometric, about 33 nm in diameter; in 2% uranyl acetate (pH 7.0) they are regular polyhedrons with angular outlines measuring about 35 nm in diameter (Fig. 6).
Particle Composition
Nucleic acid: Single-stranded RNA comprising about 20% of particle weight (estimated spectrophotometrically) (C. Hiruki, unpublished information). Two species of RNA, of M. Wt 1.35 x 106 and 0.55 x 106, determined under non-denaturing conditions, comprise the viral genome (Okuno et al., 1983).
Protein: The virus particles contain a single protein species of M. Wt 38,000, comprising about 80% of the particle weight (Hiruki et al., 1984c).
Genome Properties
Both RNA-1 and RNA-2 are essential for infectivity. RNA-1 carries determinants for the particle protein and for systemic invasion of sweet clover at 26°C. Several other types of symptom are determined by interaction between RNA-1 and RNA-2 (Okuno et al., 1983).
Relations with Cells and Tissues
Large amorphous aggregates consisting of spherical virus particles were observed in the cytoplasm of infected cells (Fig. 5). These were similar to the amorphous bodies observed in the epidermal cells of infected plants in the light microscope.
Notes
In the early literature, more than 20 viruses were reported to affect M. alba (more than 10 for M. officinalis) (Thornberry, 1966). Sweet clover necrotic mosaic virus differs from most of these viruses in possessing isometric particles c. 34 nm in diameter which sediment as a single component and contain two genomic RNA species of M. Wt about 1.4 x 106 and 0.6 x 106. The virus can easily be separated from a mixture with other viruses because of its high dilution end-point (10-6), high thermal inactivation point (90°C, 10 min), and stable infectivity (10 weeks in dried leaf tissue at room temperature), or by using certain differential hosts. Lucerne transient streak virus, recently detected in alfalfa in Canada (Rao & Hiruki, 1985a), has also been found naturally infecting sweet clover, sometimes together with sweet clover necrotic mosaic virus (C. Hiruki, unpublished data).
Figures

Sweet clover (Melilotus officinalis) leaves: (left to right), uninoculated control; inoculated leaf with ringspot and veinal necrosis symptoms; and three systemically infected leaves showing veinclearing, mosaic, and veinal necrosis.

Necrotic local lesions in Chenopodium amaranticolor.

Necrotic local lesions in Vigna unguiculata cv. Early Ramshorn.

Necrotic local lesions in Phaseolus vulgaris cv. Red Kidney.
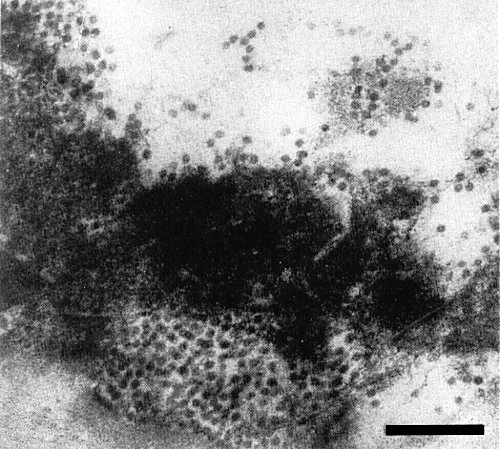
Aggregates of virus particles in a necrotic cell of Phaseolus vulgaris cv. Red Kidney 7 days after inoculation. Bar represents 200 nm.

Partially purified preparation of virus particles, stained in uranyl acetate (pH 7.0). Bar represents 100 nm.
References list for DPV: Sweet clover necrotic mosaic virus (321)
- Chen, Hiruki & Okuno, Can. J. Pl. Path. 6: 191, 1984.
- Gould, Francki, Hatta & Hollings, Virology 108: 499, 1981.
- Hiruki, Proc. 4th int. Conf. Compar. Virol., Banff, Canada, p. 57, 1982.
- Hiruki, Pl. Dis. 70: 1129, 1986.
- Hiruki & Figueiredo, Acta Hort. 164: 217, 1985.
- Hiruki, Okuno, Rao & Chen, Abstr. 5th int. Congr. Virol., Strasbourg, France, p. 235, 1981.
- Hiruki, Figueiredo, Inouye & Furuya, J. virol. Meth. 8: 301, 1984a.
- Hiruki, Rao, Furuya & Figueiredo, J. gen. Virol. 65: 2273, 1984b.
- Hiruki, Rao, Chen, Okuno & Figueiredo, Phytopathology 74: 482, 1984c.
- Inouye & Hiruki, Ann. phytopath. Soc. Japan 51: 82, 1985.
- Lommel, Ph.D. Thesis, Univ. Calif., Berkeley, 1983.
- Matthews, Intervirology 17: 1, 1982.
- Okuno, Hiruki, Rao & Figueiredo, J. gen. Virol. 64: 1907, 1983.
- Rao & Hiruki, Pl. Dis. 69: 610, 1985a.
- Rao & Hiruki, Pl. Dis. 69: 959, 1985b.
- Thornberry, Agriculture Handbook No. 307, U.S. Dept. Agric., 1966.