Details of DPV and References
DPV NO: 324 December 1986
Family: Potyviridae
Genus: Potyvirus
Species: Iris mild mosaic virus | Acronym: IMMV
This is a revised version of DPV 116
Iris mild mosaic virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, West Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described by Van Slogteren (1958, 1960) and Loebenstein & Alper (1963); virus partially characterized by Brunt & Phillips (1980), Inouye et al. (1981) and Derks et al. (1985)
Selected synonyms
- Iris mosaic virus (Rev. appl. Mycol 46: 350; 47: 2183), but now obsolete.
- Iris latent mosaic virus (Rev. appl. Mycol 43: 881e).
-
A virus with filamentous particles c. 750 x 12 nm, prevalent in bulbous iris but reported also in rhizomatous iris and Ferraria undulata. Transmissible in the non-persistent manner by aphids, and by inoculation of sap to monocotyledonous and dicotyledonous species. World-wide distribution.
Main Diseases
When occurring alone, the virus usually causes mild leaf chlorosis in field-grown bulbous irises, especially in I. x hollandica (Dutch iris), I. anglica (English iris), I. xiphium (Spanish iris), I. histrioides, I. danfordiae and I. reticulata (Van Slogteren, 1958, 1960; Asjes, 1979); however, it frequently occurs together with iris severe mosaic, bean yellow mosaic and/or narcissus latent viruses, in which event irises are more severely affected. Iris mild mosaic virus has been detected also in the rhizomatous I. susiana (O. W. Barnett & M. Alper, personal communication), and in Ferraria undulata (Iridaceae) with light and dark green mottling of the leaves (Derks et al., 1985).
Geographical Distribution
World-wide; older stocks of commercially important cultivars of bulbous iris are probably totally infected (Van Slogteren, 1958; Loebenstein & Alper, 1963; Lawson, 1967; Asjes, 1979), but virus-free stocks are now available in some countries including Britain and The Netherlands (Asjes, 1983; Brunt, 1985; Derks, 1985).
Host Range and Symptomatology
In addition to its natural iridaceous hosts, the virus is reported to infect 16 species in five dicotyledonous families (Brunt & Phillips, 1980; Inouye et al., 1981; Franco & Russo, 1981; Derks et al., 1985).
-
Diagnostic species
- Chenopodium quinoa.
Many isolates induce numerous circular chlorotic lesions in inoculated leaves (Fig. 3); no systemic infection. - Freesia refracta cv. Imperial. Small dark green spots and chlorosis of systemically
infected leaves (Fig. 2) and dark blue spots in petals.
- Iris x hollandica cvs Wedgwood and Professor Blaauw. Chlorotic stripes in inoculated leaves of glasshouse-grown plants after 2-4 weeks, soon followed by mottling of systemically infected leaves (Fig. 1). Symptoms in glasshouse-grown plants are initially more severe than those induced under similar conditions by iris severe mosaic virus (Brunt & Phillips, 1980; Derks et al., 1985).
-
Propagation species
- I.
x hollandica and I. reticulata are suitable for maintenance of cultures. Inoculated leaves of Chenopodium quinoa are a good source of some isolates for virus purification.Assay species
- Chenopodium quinoa
is a sensitive local lesion assay host for many isolates.
Strains
None distinguished.
Transmission by Vectors
Transmissible in the non-persistent manner by Aphis gossypii, Macrosiphum euphorbiae amd Myzus persicae, but not by A. craccivora. Aphids acquire and inoculate virus in 5-10 min feeding periods; no latent period (Loebenstein & Alper, 1963; Inouye et al., 1981; S. Phillips & A. A. Brunt, unpublished information; A. F. L. M. Derks, unpublished information).
Transmission through Seed
Not seed-borne in Iris x hollandica.
Serology
The virus is a good immunogen. Antiserum with a homologous titre of 1/640 in microprecipitin tests has long been available in the Netherlands (Van Slogteren, 1955; see Asjes, 1979 and Derks et al., 1980). Antiserum with a homologous titre of 1/16,000-1/32,000 in tube precipitin tests was obtained from a rabbit 4 weeks after the second of two intramuscular injections (administered 2 weeks after the first) of c. 0.75 mg virus emulsified with Freund's complete adjuvant (A. A. Brunt & S. Phillips, unpublished information). An antiserum which is particularly suitable for ELISA has been produced in Israel (M. Alper & G. Loebenstein, unpublished information).
Relationships
Although a typical member of the potyvirus group, the virus showed no serological relationship in tube precipitin tests to iris severe mosaic (Van Slogteren, 1958; Loebenstein & Alper, 1963; Lawson, 1967), potato Y, bean yellow mosaic, turnip mosaic, alstroemeria mosaic, hippeastrum mosaic or four other morphologically similar viruses (Brunt & Phillips, 1980; Inouye et al., 1981). In indirect ELISA, however, iris mild mosaic virus was found to be related to asparagus 1, bean yellow mosaic, pea seed-borne mosaic, potato Y and tobacco etch viruses (J. Hammond, personal communication). In addition, four of eight monoclonal antibodies to tulip breaking virus (Hsu et al., 1983) reacted with iris mild mosaic virus in sap or purified preparations (Hammond et al., 1985a). Although none of the monoclonal antibodies prepared to a mixture of iris mild mosaic, iris severe mosaic and bean yellow mosaic viruses reacted solely with iris mild mosaic virus, selected antibodies can be used to distinguish the three viruses in mixed infections in iris (Hammond et al., 1985a; R. Jordan & J. Hammond, unpublished information).
Stability in Sap
In sap of Iris x hollandica cv. Professor Blaauw, the thermal inactivation point (10 min) is 65-70°C, dilution end-point 10-3 to 10-4, and infectivity is retained for 3-4 days at 20°C or 16-32 days at 2°C (Inouye et al., 1981; S. Phillips & A. A. Brunt, unpublished information).
Purification
1. (A. F. L. M. Derks, unpublished information). Homogenize infected iris leaves in either 0.2 M borate buffer at pH 7.2 or 0.067 M phosphate buffer at pH 7.2 containing 0.1% (v/v) thioglycollic acid (1 ml/g tissue). To fresh or thawed homogenate add a 1:1 mixture of chloroform and n-butanol (1 ml/g leaf tissue) and, after 30 min centrifuge at low speed. Recover the virus particles from the aqueous phase by centrifugation at 100,000 g for 2 h (Asjes, 1979). Virus can be further purified by precipitation with polyethylene glycol.
2. (S. Phillips & A. A. Brunt, unpublished information). Homogenize infected iris or Chenopodium quinoa leaves in 0.067 M phosphate buffer at pH 7.6 containing 0.1% thioglycollic acid (2 ml/g tissue). Add sulphite-washed n-butanol to 8% (v/v) and, after stirring the mixture gently for 30-60 min, concentrate the virus by one or more cycles of differential centrifugation (20 min at 10,000 g; 90 min at 90,000 g). The virus can be further purified by chromatography on controlled-pore glass beads.
3. (J. Hammond, personal communication). Homogenize infected iris leaves in 0.5 M phosphate buffer at pH 8.4 (3 ml/g leaf tissue) centrifuge the extract for 10 min at 7000 g, stir the supernatant fluid for 15 min with 2% (v/v) Triton X-100, and then for 1 h after adding polyethylene glycol (M. Wt 20,000) to 4% (w/v) and sodium chloride to 0.1 M. Centrifuge the mixture for 10 min at 7000 g, resuspend the precipitate in 1/5th the original volume of 0.1 M borate buffer at pH 8.0, clarify by centrifugation (10 min at 7000 g) and sediment the virus particles from the supernatant fluid through a 30% sucrose cushion for 2.5 h at 62,900 g. Resuspend the sedimented virus in a small volume of 0.1 M borate buffer, add caesium chloride to 1.32 g cm-3, centrifuge for 16-20 h at 103,000 g, remove the virus-containing band and dialyse against 50 mM borate buffer at pH 8.0.
Properties of Particles
Purified virus particles usually sediment as a single component with a sedimentation coefficient (s°20, w) of 142-147 S.
A260(max)/A242(min): 1.1.
A260/A280: 1.18-1.22.
Buoyant density in CsCl: 1.32 g cm-3.
Particle Structure
The particles (Fig. 4) are slightly flexuous filaments mostly measuring c. 750 x 12 nm (Asjes, 1979; Brunt & Phillips, 1980; Inouye et al., 1981; Derks et al., 1985).
Particle Composition
Nucleic acid: RNA c. 5% of particle weight (estimated spectrophotometrically).
Protein: c. 95% of particle weight; in SDS-polyacrylamide gels the particle protein migrated as a single band with an estimated M. Wt of 33 x 103 (Alper et al., 1984; Hammond & Lawson, 1985).
Relations with Cells and Tissues
Aggregates of virus particles and cylindrical inclusions are found in the cytoplasm of
infected iris and Chenopodium quinoa cells
(Fig. 5, Fig. 6, Fig. 7);
the inclusions are seen in section
primarily as pinwheels with laminated aggregates, the inclusion lamellae occurring in close
association with virus particles (J. R. Edwardson, personal communication;
Inouye et al., 1981;
M. Alper & O. W. Barnett, personal communication). Fragments of pinwheel lamellae
present in plant extracts are often triangular with a lattice of 5.3 nm
(Inouye et al., 1981).
The inclusions consist of a single protein of M. Wt 69 x 103
(Alper et al., 1984;
Hammond & Lawson, 1985).
Iris mild mosaic, bean yellow mosaic and
iris severe mosaic viruses
can be distinguished, especially when occurring in complex in iris, by differences
in the apparent M. Wt of their inclusion proteins
(Alper et al., 1984;
Hammond & Lawson, 1985).
Nuclei of infected plants (Fig. 8) also sometimes contain fibrous inclusions (J. R. Edwardson, personal communication; M. Alper & O. W. Barnett, unpublished information).
Notes
Two distinct aphid-borne potyviruses have long been known to occur prevalently in commercial stocks of bulbous iris (Brierley & McWhorter, 1936; Van Slogteren, 1958, 1960; Loebenstein & Alper, 1963). Although the nomenclature of the two viruses has previously been confused, confusion can best be avoided by designating them iris mild mosaic and iris severe mosaic viruses (Lawson, 1967; Hammond et al., 1985b). Although the two viruses are morphologically indistinguishable, they are serologically distinct and have different experimental host ranges; thus, iris mild mosaic virus induces conspicuous symptoms both in monocotyledonous and in dicotyledonous species whereas iris severe mosaic virus apparently infects only monocotyledonous species and, unlike the milder virus, induces conspicuous chlorosis and/or mosaic patterns in systemically infected leaves of Belamcanda chinensis and Crocus vernus (Derks et al., 1985).
Iris mild mosaic virus also differs from the potyvirus
iris fulva mosaic virus, found
infecting only I. fulva hybrids in north eastern USA; the two viruses are serologically
distinct, and RNA from iris mild mosaic virus fails to hybridize with cDNA to iris fulva
mosaic virus RNA
(Barnett, 1986). Moreover, although the cytoplasmic inclusions induced by
both viruses are usually seen in leaf sections as pinwheels and laminated aggregates, only
with iris mild mosaic virus are particles usually associated with the inclusion lamellae.
Other viruses also sometimes occur in bulbous irises containing iris mild mosaic and iris
severe mosaic viruses. Of the three transmitted by aphids,
bean yellow mosaic virus has been
reported from England, Israel, The Netherlands and USA
(Brunt & Phillips, 1980;
Derks et al., 1980;
Alper & Loebenstein, 1981;
Brierley & Smith, 1948;
Hammond & Lawson, 1985),
turnip mosaic virus from Japan
(Inouye & Mitsuhata, 1978) and
narcissus latent virus from England and The Netherlands
(Brunt, 1977;
Asjes, 1979).
Tobacco ringspot,
tobacco mosaic and
tobacco rattle viruses have been detected occasionally in bulbous iris in
The Netherlands
(Asjes, 1969, 1974). These six viruses, however, have been well characterized
and are readily distinguishable from iris mild mosaic and iris severe mosaic viruses by host
range, symptomatology and other properties.
Iris mild mosaic virus is now so prevalent that virus-free stocks of commercially important
cultivars are probably unobtainable by selection and roguing. Although the virus cannot be
eliminated from bulbs by heat treatment
(Kassanis & Broadbent, 1958), virus-free plants
are obtainable by meristem-tip culture
(e.g. Stone & Hollings, 1965;
Baruch & Quak, 1966;
Brunt, 1985;
Derks, 1985).
Acknowledgements
Figs 1 and 2 courtesy of A. F. L. M. Derks, Figs 5, 6 and 7 courtesy of N. Inouye and Fig. 8 courtesy of M. Alper and O. W. Barnett.
Figures

Chlorotic striping in systemically infected leaves of Iris x hollandica cv. Wedgwood.

Healthy (left) and infected (right) leaves of Freesia refracta cv. Imperial.

Lesions induced by iris mild mosaic virus in inoculated leaves of Chenopodium quinoa.

Particles from a purified preparation mounted in neutral phosphotungstate. Bar represents 250 nm.

Aggregates of virus particles in infected C. quinoa leaf. Bar represents 500 nm.

Cytoplasmic inclusions, laminated aggregates and virus particles closely associated with pinwheel lamellae. Bar represents 500 nm.

Pinwheel lamellae with associated virus particles. Bar represents 250 nm.
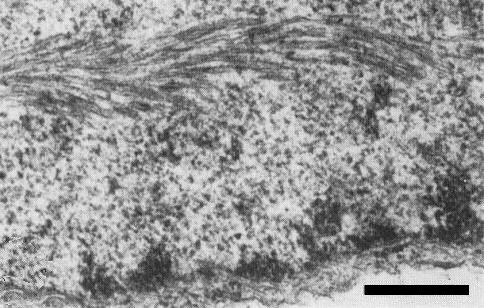
Fibrous nuclear inclusion. Bar represents 250 nm.
References list for DPV: Iris mild mosaic virus (324)
- Alper & Loebenstein, Pl. Dis. 65: 694, 1981.
- Alper, Salomon & Loebenstein, Phytopathology 74: 960, 1984.
- Asjes, Jversl. Lab. BloembollenOnderz. Lisse, 1968-69: 44, 1969.
- Asjes, Agric. Envir. 1: 303, 1974.
- Asjes, Neth. J. Pl. Path. 85: 269, 1979.
- Asjes, Bloembollencultuur 93: 51 & 77, 1983.
- Barnett, AAB Descr. Pl. Viruses 310, 4 pp., 1986.
- Baruch & Quak, Neth. J. Pl. Path. 72: 270, 1966.
- Brierley & McWhorter, J. agric. Res. 53: 621, 1936.
- Brierley & Smith, Phytopathology 38: 574, 1948.
- Brunt, Ann. appl. Biol. 87: 355, 1977.
- Brunt, Acta Hort. 164: 153, 1985.
- Brunt & Phillips, Acta Hort. 109: 503, 1980.
- Derks, Acta Hort. 164: 281, 1985.
- Derks, Vink-van den Abeele & Muller, Acta Hort. 110: 31, 1980.
- Derks, Hollinger & Vink-van den Abeele, Acta Hort. 164: 309, 1985.
- Franco & Russo, Riv. Ortoflorofruttic. ital. 65: 301, 1981.
- Hammond & Lawson, Acta Hort. 164: 225, 1985.
- Hammond, Lawson & Hsu, Phytopathology 75: 1353, 1985a.
- Hammond, Brunt, Derks, Inouye, Barnett, Allen & Lawson, Acta Hort. 164: 395, 1985b.
- Hsu, Lawson, Beijersbergen & Derks, Proc. 4th int. Congr. Pl. Path. p. 117, 1983.
- Inouye & Mitsuhata, Nogaku Kenkyu 57: 1, 1978.
- Inouye, Miyaji & Mitsuhata, Ann. phytopath. Soc. Japan 47: 182, 1981.
- Kassanis & Broadbent, Rep. Rothamsted exp. Stn, 1957: 106, 1958.
- Lawson, Pl. Dis. Reptr 51: 952, 1967.
- Loebenstein & Alper, Phytopathology 53: 349, 1963.
- Stone & Hollings, Rep. Glasshouse Crops Res. Inst., 1964: 95, 1965.
- Van Slogteren, Ann. appl. Biol. 42: 122, 1955.
- Van Slogteren, Jversl. Lab. BloembollenOnderz. Lisse, 1957: 66, 1958.
- Van Slogteren. Jversl. Lab. BloembollenOnderz. Lisse, 1959: 37, 1960.