Details of DPV and References
DPV NO: 325 September 1988
Family: Potyviridae
Genus: Potyvirus
Species: Tobacco vein mottling virus | Acronym: TVMV
Tobacco vein mottling virus
T. P. Pirone Department of Plant Pathology, University of Kentucky, Lexington, Kentucky 40546, USA
J. G. Shaw Department of Plant Pathology, University of Kentucky, Lexington, Kentucky 40546, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described by Gooding & Sun (1972). Virus characterized by Pirone et al. (1973) and Sun et al. (1974).
A virus with flexuous filamentous particles with a modal length of 765 nm and a diameter of 13 nm, containing single-stranded RNA. The virus induces the formation of cylindrical (pinwheel) inclusions in the cytoplasm of host cells. It is readily transmitted mechanically, and by several species of aphid in the non-persistent manner.
Main Diseases
Other than tomato, in which the virus causes symptomless infection in most cultivars, the only known host of economic importance is tobacco (Nicotiana tabacum) (Sun et al., 1974; T. P. Pirone, unpublished data). The disease is particularly severe on the Burley types grown in the southeastern USA (Pirone et al., 1973). The characteristic symptom is the presence of discontinuous blotches along the veins (Fig. 1 and Fig. 2) which can be distinguished from the continuous vein banding caused by potato virus Y. As plants mature, necrosis of veinal and sometimes of laminar tissue occurs, leading to loss of leaves; the severity of this symptom varies with the cultivar (Pirone & Gooding, 1973).
Geographical Distribution
The known distribution of the virus is limited to the Burley tobacco-producing States of the southeastern USA. A virus which causes similar symptoms on tobacco has been reported from Nigeria (Ladipo & Roberts, 1979); its possible relationship to tobacco vein mottling virus is discussed under Notes.
Host Range and Symptomatology
The virus has a very narrow host range, apparently limited to the Solanaceae (Pirone et al., 1973; Sun et al., 1974). Although infection of Rumex sp. has been reported (Sun et al., 1974), further testing has not confirmed this (G. V. Gooding & T. P. Pirone, unpublished data).
-
Diagnostic species
- Nicotiana tabacum
(tobacco). In Burley cultivars such as Kentucky 14 or Burley 21, the initial systemic symptoms are a very mild vein-clearing with occasional blotches along the veins (Fig. 1); subsequent symptoms are a pronounced veinal blotching or mottling (Fig. 2). Symptoms are much less distinct on other types of tobacco such as Xanthi, Samsun and Havana 425. The type strain induces no symptoms on the ‘Virgin A’ mutant (TI 1406) and virus can be recovered only from inoculated leaves (Johnson & Pirone, 1982). - Lycopersicon esculentum (tomato). The cv. Rutgers is a symptomless host (T. P. Pirone,
unpublished data); mild mottling is produced in cv. Homestead
(Sun et al., 1974).
- Capsicum annuum (sweet pepper) and C. frutescens (Tabasco pepper). No symptoms are produced, nor can virus be recovered, from plants inoculated mechanically or by aphids (Sun et al., 1974; T. P. Pirone, unpublished data).
-
Propagation species
- Cultivars of Burley tobacco (e.g., Kentucky 14 or Burley 21) which possess hypersensitive resistance to tobacco mosaic virus are good hosts for virus propagation and purification.
-
Assay species
- No local lesion hosts for the virus have been reported. Kentucky 14 tobacco is useful for systemic assays.
Strains
As with other potyviruses, the distinction between strains and other named viruses is somewhat arbitrary (see Relationships). Twenty-five naturally occurring isolates from tobacco could not be differentiated by the symptoms induced in the tobacco cultivars Kentucky 14, Burley 21 or Kentucky 14 x L8, or serologically using the agar-gel double-diffusion technique (Gooding & Rufty, 1987). However, isolates of tobacco vein mottling virus which infect the ‘Virgin A’ mutant tobacco (TI 1406) systemically, and thus differ from the type strain, have been reported (Johnson, 1980).
Transmission by Vectors
Transmitted in the non-persistent manner by aphids. Acquisition and inoculation thresholds are about 15-30 sec, and the virus is retained from minutes to hours, depending upon aphid behaviour (T. P. Pirone, unpublished data). Reported vectors are Myzus persicae, Macrosiphum euphorbiae, Aphis craccivora, A. gossypii, Rhopalosiphum maidis and R. padi (Pirone et al., 1973; Kennedy et al., 1979). By analogy with other potyviruses, it is likely that other species of aphid are capable of transmitting the virus.
The presence of ‘helper component’, a virus-encoded protein, is required for transmission of the purified virus by aphids (Pirone, 1981). In the presence of helper component, purified virus acquired through membranes is retained in the food canal or other parts of the anterior alimentary tract, whereas if helper component is not present, virus passes into the gut (Berger & Pirone, 1986).
As with other potyviruses, serial transmission by mechanical inoculation can lead to the development of isolates which are not aphid-transmissible (D. W. Thornbury & T. P. Pirone, unpublished data).
Transmission through Seed
None reported.
Serology
The virus is a good immunogen. Antisera with titres of 1/1024 in ring-precipitin tests have been prepared. Immunodiffusion tests in SDS agar (Gooding & Bing, 1970) can be used for detection of virus in crude sap extracts, and both direct and indirect ELISA are useful for detection and quantitation of the virus (K. S. Gibb & T. P. Pirone, unpublished data).
Anti-helper component antisera have been used in neutralization of activity tests which distinguish the helper components of tobacco vein mottling virus and potato virus Y (Thornbury & Pirone, 1983). These antisera have been used to immunoprecipitate the products of cell-free translation of the RNA molecules of several potyviruses, and so to demonstrate the presence and relationships of helper component proteins (Hiebert et al., 1984).
Relationships
The chemical and physical properties of the particles, aphid transmissibility, and the production of pinwheel inclusions in host cells place the virus in the potyvirus group. In SDS-agar double-diffusion tests the virus failed to react with antisera to henbane mosaic, peanut mottle, soybean mosaic, iris severe mosaic (bearded iris mosaic), bean common mosaic and clover yellow vein viruses (Sun et al., 1974).
Because of the limited host range most of the detailed serological comparisons have been made with potyviruses that primarily infect Solanaceae. Although the virus and its antiserum failed to react with antisera and antigens of potato virus Y or tobacco etch virus in SDS-agar diffusion tests using crude sap extracts (Sun et al., 1974; see also Fig. 5), reciprocal ring-precipitin tests with purified viruses show a moderately distant relationship with tobacco etch virus (SDI 2-3) and a distant relationship with potato virus Y (SDI 5-6) (T. P. Pirone, unpublished data). The virus has been reported to be closely related to the type and the Nigerian tobacco virus strains of pepper veinal mottle virus by electron microscope serology (Ladipo & Roberts, 1979), but ring precipitin and ELISA tests indicate a distant relationship (T. P. Pirone, unpublished data).
Stability in Sap
In tobacco sap, the thermal inactivation point is between 60 and 70°C, dilution end-point between 10-2 and 10-3, and longevity in vitro about 2 days at 22-25°C (Sun et al., 1974). Infectivity survives in desiccated leaf tissue at 4°C for several years (T. P. Pirone, unpublished data).
Purification
A modification of a method reported by Reddick & Barnett (1983) is rapid and effective (J. F. Murphy, personal communication). In a blender, briefly grind 100 g systemically infected tobacco leaves in 150 ml cold 0.5 M phosphate buffer, pH 7, containing 1 M urea and 0.1% 2-mercaptoethanol. Add 50 ml cold CHCl3 and 50 ml cold CCl4 and homogenize for 1 min. Centrifuge the material at 8000 g for 15 min and filter the aqueous (upper) phase through glass wool. To the filtrate, add NaCl to 0.25 M and PEG 8000 to 4% (w/v) and stir the mixture for 1 h at 4°C. Centrifuge at 8000 g for 15 min and resuspend the pellets in 50-100 ml 0.1 M phosphate buffer, pH 7, containing 1 M urea. Add Triton X-100 to 1% and stir for 2 h at 4°C. Centrifuge and treat the supernatant fluid with NaCl and PEG as described above. Resuspend the pellet in 8 ml 0.02 M Tris-HCl, pH 7.5, and add 1.6 g Cs2SO4. Layer 4.2 ml of suspension over 0.8 ml 53% (w/w) Cs2SO4 in each of two Beckman SW50.1 rotor tubes. Centrifuge for 16-18 h at 32,000 rev./min at 4°C. Collect the virus, dilute with buffer and concentrate by centrifugation at 45,000 rev./min for 1 h in the Beckman 65 rotor. Yields of c. 10 mg virus/100 g tissue are normal. The approximate concentration of the virus can be estimated spectrophotometrically using A0.1%, 260 nm = 2.4.
Properties of Particles
Sedimentation coefficient, buoyant density in caesium salts and extinction coefficient at 260 nm have not been determined but are presumed to be similar to those of other potyviruses.
Particle Structure
The particles are flexuous filaments c. 765 nm long with a diameter of 13 nm (Sun et al., 1974 and Fig. 3).
Particle Composition
Nucleic Acid: Single-stranded RNA of M. Wt c. 3.2 x 106. One molecule per particle, presumed to be c. 5% of particle weight. The nucleotide sequence (9471 bases) has been determined (Domier et al., 1986).
Coat Protein: Approximately 29,000 M. Wt. Number of molecules per particle and details of their arrangement in the particle are not known. The protein undergoes partial proteolysis during storage.
Genome Properties
The monopartite, positive-sense RNA carries a genome-linked protein (VPg) at the 5'-terminus (Siaw et al., 1985) and poly(A) at the 3' terminus. The M. Wt of the VPg has been reported as c. 24,000 (Siaw et al., 1985) but is probably much smaller.
The RNA contains one long open reading frame (ORF), beginning 206 nucleotide residues from the 5'-terminus, which could encode a polyprotein of M. Wt 340,000 (Fig. 6) (Hellmann et al., 1986). The ORF contains cistrons for five known proteins: helper component (HC), cylindrical inclusion protein (CI), two nuclear inclusion proteins (NIa and NIb) and coat protein (CP), and coding capacity for three additional polypeptides of M. Wt (x 10-3) c. 28, 42 and 5·5. The VPg is encoded by the 5'-portion of the NIa cistron (Shahabuddin et al., 1988); NIa functions as a protease which cleaves the polyprotein at several sites to yield some of the mature proteins (Hellmann et al., 1988). An additional virus-encoded protease may exist. CI, NIa and NIb have been shown to share amino acid sequence homology with proteins considered to be involved in RNA replication of comoviruses and picornaviruses (Domier et al., 1987).
The genome of tobacco vein mottling virus is probably expressed in vivo by proteolytic cleavage of the polyprotein. There is no evidence of the presence of subgenomic mRNA species in infected cells (Luciano et al., 1987). In vitro translation of the genomic RNA in the rabbit reticulocyte lysate system yields a complex array of polypeptides (Hellmann et al., 1980, 1983).
Relations with Cells and Tissues
Pinwheel inclusions, scrolls and laminated aggregates are produced in the cytoplasm of infected cells (Fig. 4).
Notes
Symptoms produced by tobacco vein mottling virus on field-grown tobacco may be confused with those produced by tobacco etch virus or potato virus Y, both of which commonly occur in the same geographical area. The viruses are readily differentiated by serology in SDS-agar diffusion tests (Fig. 5). The limited host range of tobacco vein mottling virus also distinguishes it from these viruses. Specific indicator plants such as Tabasco pepper, in which only tobacco etch virus causes wilt, and Solanum demissum, which is susceptible to potato virus Y but not the other two viruses, are useful differential hosts.
A strain of pepper veinal mottle virus from Nigeria, which causes symptoms in tobacco identical to those produced by tobacco vein mottling virus, was reported by Ladipo & Roberts (1979). The host range of the Nigerian tobacco isolate is more extensive than that of tobacco vein mottling virus; in particular, peppers (Capsicum annuum and C. frutescens), which are immune to tobacco vein mottling virus, are readily infected with the Nigerian tobacco isolate (Ladipo & Roberts, 1979). Although both the type and Nigerian tobacco isolates of pepper veinal mottle virus were found to be related to tobacco vein mottling virus by electron microscope serology (Ladipo & Roberts, 1979), ring-precipitin tests and ELISA showed only a distant relationship (T. P. Pirone, unpublished data).
Figures

Vein clearing and vein blotching on the first systemically infected leaf of tobacco cultivar Kentucky 14.

Characteristic mottling and blotching produced along the veins of Burley 21 tobacco systemically infected with the virus.
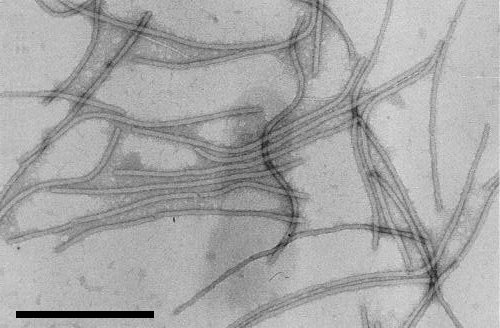
Purified virus particles, stained in uranyl acetate. Bar represents 400 nm. (Courtesy U. Jarlfors.)

Ultrathin section showing cytoplasmic inclusions in infected tobacco cells. Bar represents 375 nm. (Courtesy U. Jarlfors.)

Sodium dodecyl sulphate immunodiffusion tests with sap extracts of tobacco infected with tobacco etch virus (E), tobacco vein mottling virus (T), potato virus Y (Y) and antiserum to particles of tobacco vein mottling virus (A).

Proposed genome map for tobacco vein mottling virus. Known gene products are helper component (HC), cylindrical inclusion protein (CI), the genome linked protein (VPg), nuclear inclusion proteins (NIa and NIb) and the coat protein (CP). The approximate molecular weights of three additional putative gene products are also indicated.
References list for DPV: Tobacco vein mottling virus (325)
- Berger & Pirone, Virology 153: 256, 1986.
- Domier, Franklin, Shahabuddin, Hellmann, Overmeyer, Hiremath, Siaw, Lomonossoff, Shaw & Rhoads, Nucl. Acids Res. 14: 5417, 1986.
- Domier, Shaw & Rhoads, Virology 158: 20, 1987.
- Gooding & Bing, Phytopathology 60: 1293, 1970.
- Gooding & Rufty, Plant Dis. 71: 38, 1987.
- Gooding & Sun, Phytopathology 62: 803, 1972.
- Hellmann, Shaw, Lesnaw, Chiu, Pirone & Rhoads, Virology 106: 207, 1980.
- Hellmann, Thornbury, Hiebert, Shaw, Pirone & Rhoads, Virology 124: 434, 1983.
- Hellmann, Hiremath, Shaw & Rhoads, Virology 151: 159, 1986.
- Hellmann, Shaw & Rhoads, Virology 163: 554, 1988.
- Hiebert, Thornbury & Pirone, Virology 135: 1, 1984.
- Johnson, Ph.D. Thesis, University of Kentucky, Lexington, 1980.
- Johnson & Pirone, Phytopathology 72: 68, 1982.
- Kennedy, Wade & Gooding, Phytopathology 69: 529, 1979.
- Ladipo & Roberts, Pl. Dis. Reptr 63: 161, 1979.
- Luciano, Rhoads & Shaw, Pl. Sci. 51: 295, 1987.
- Pirone, Phytopathology 71: 922, 1981.
- Pirone & Gooding, Pl. Dis. Reptr 57: 845, 1973.
- Pirone, Gooding & Smiley, Pl. Dis. Reptr 57: 841, 1973.
- Reddick & Barnett, Phytopathology 73: 1506, 1983.
- Shahabuddin, Shaw & Rhoads, Virology 163: 635, 1988.
- Siaw, Shahabuddin, Ballard, Shaw & Rhoads, Virology 142: 134, 1985.
- Sun, Gooding, Pirone & Tolin, Phytopathology 64: 1133, 1974.
- Thornbury & Pirone, Virology 125: 487, 1983.