Details of DPV and References
DPV NO: 326 September 1988
Family: Pospiviroidae
Genus: Hostuviroid
Species: Hop stunt viroid | Acronym: HSVd
Hop stunt viroid
T. Sano Department of Botany, Faculty of Agriculture, Hokkaido University, Sapporo 060, Japan
E. Shikata Department of Botany, Faculty of Agriculture, Hokkaido University, Sapporo 060, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Disease first described by Yamamoto et al. (1970), the viroid by Sasaki & Shikata (1977a, 1977b).
- Synonyms
Grapevine viroid (Shikata et al., 1984)
Cucumber pale fruit viroid (disease observed since 1965, viroid described by Van Dorst & Peters, 1974) is closely related, though not identical, to isolates from hop and grapevine.
A covalently closed circular or single-stranded linear RNA occurring in almost all hop gardens in the northern part of the mainland of Japan. The form in hop is detected only in hop but closely related viroids occur in grapevine (‘grapevine viroid’) and cucumber (‘cucumber pale fruit viroid’) and are treated here as strains. The hop form is mechanically transmissible from nucleic acid inocula and from hop to hop by normal cultural practices. The cucumber form, found only in glasshouse-grown cucumber in the Netherlands, is transmitted during pruning.
Main Diseases
Causes stunt, a serious disease of hop (Humulus lupulus) recognized by dwarfing of the plant, shortening of the internodes of the main and lateral bines, yellowing and curling of the upper leaves. The main bines of diseased hops reach only c. 5 m, 60% of the height of healthy ones (Fig. 1). The cones of diseased hop are smaller (Fig. 2), and the number per bine fewer (c. 50%), than those of healthy plants, resulting in about 50% loss in cone yield. The a-acid content in the bitter substances in cones is only one half to one third of that in cones from healthy plants (Yamamoto et al., 1970, 1973).
The cucumber strain induces pale fruits, crumpled flowers, and rugosity and chlorosis of the leaves of cucumber grown in glasshouses. The disease spreads slowly, but the number of affected plants is usually low, mostly less than 0.1% (Van Dorst & Peters, 1974).
The effects of the grapevine strain on grapevine are unknown.
Geographical Distribution
In Japan, hop is cultivated primarily in Tohoku District (northern part of the mainland, Honshu), Nagano and Hokkaido. Hop stunt disease is prevalent in almost all hop gardens in Tohoku and Nagano (Yamamoto et al., 1970, 1973; Sasaki & Shikata, 1980). Cucumber pale fruit disease is reported only in The Netherlands, where it occurs in glasshouses throughout the country (Van Dorst & Peters, 1974). The grapevine strain can be detected in many grapevine cultivars with high frequency (c. 85%), and is distributed world-wide (Europe, USA, China and Japan) (Sano et al., 1986).
Host Range and Symptomatology
The hop, cucumber and grapevine strains can be transmitted mechanically to several species of Cucurbitaceae, Solanaceae and Moraceae (Sasaki & Shikata, 1977a; Van Dorst & Peters, 1974; Sano et al., 1986). The host ranges and symptoms of the three strains are similar (Sano et al., 1981, 1985).
-
Diagnostic species
- Benincasa hispida
(white gourd). Infected plants show stunting, mosaic, leaf curling and tip necrosis. Some severely infected plants die. - Cucumis sativus (cucumber) cv. Suyo. Plants infected by rub-inoculation of crude sap on fully expanded cotyledons show stunting, vein clearing and downward leaf curling after 14-28 days at 25-32°C (Fig. 3, Fig. 4).
-
Propagation species
- Cucumis sativus
cv. Suyo.- Assay species
- No local lesion assay host is known. The viroid may be assayed by determining the proportion of Cucumis sativus cv. Suyo plants that become infected. After inoculation, plants should be maintained at 25-32°C, 16 h daylength, for at least 6 weeks.
Strains
The host range and symptoms of hop stunt viroid are similar to those of grapevine and cucumber pale fruit viroids (Van Dorst & Peters, 1974; Sasaki & Shikata, 1977a, 1978; Sano et al., 1981, 1985) and, though the nucleotide sequences of the three viroids differ slightly, their homology is extremely high (95% to 99%) (Sano et al., 1984, 1985). These properties suggest that the three viroids are closely related, and support the designation of cucumber pale fruit viroid and grapevine viroid as cucumber and grapevine strains of hop stunt viroid. No isolates of the hop strain differing in the symptoms induced in field-grown hops have been recognized. Synthetic oligonucleotide probes (e.g. 5'GGTAAGTACCTCCCT3'; complementary to residues 51-65 of the grapevine strain) could be used to discriminate between the three strains in dot-blot hybridization tests (T. Sano et al., 1988).
Transmission by Vectors
No vector has been reported. Tests for transmission of the hop and cucumber strains through soil or by aphids (Myzus persicae) were negative (Sasaki & Shikata, 1978; Van Dorst & Peters, 1974; Yamamoto et al., 1970, 1973).
The hop strain is transmissible by hands, sickles, scissors and bine contact during dressing, pulling and picking in the fields, and experimentally by razor blades (Sasaki & Shikata, 1978, 1980; Takahashi & Yaguchi, 1985).
Transmission through Seed
No seed transmission was detected of the hop strain in hop and Lycopersicon esculentum (tomato) cvs. Rutgers and Rentita, or of the cucumber strain in cucumber (Sasaki & Shikata, 1978; Sano et al., 1981; Yaguchi & Takahashi, 1984; Van Dorst & Peters, 1974).
Transmission by Dodder
No information is available for the hop or grapevine strains. The cucumber strain can be transmitted by dodder (Cuscuta subinclusa), but not by C. campestris (Van Dorst & Peters, 1974).
Relationships
Hop stunt viroid is different from seven other viroids (avocado sunblotch, chrysanthemum stunt, citrus exocortis, coconut cadang-cadang, potato spindle tuber, tomato apical stunt and tomato planta macho) in host range, symptoms, molecular size, electrophoretic mobility in polyacrylamide gels (Fig. 5) and nucleotide sequence. Hop stunt viroid causes symptoms mainly in hop and cucurbitaceous plants (e.g. cucumber) and infects species of Solanaceae (e.g. tomato) symptomlessly; in contrast, viroids related to potato spindle tuber viroid cause symptoms mainly in species of Solanaceae (e.g. tomato) and Compositae (e.g. Gynura) but infect symptomlessly or do not infect cucurbitaceous plants. Hop stunt viroid has 297 nucleotides (hop and grapevine strains) or 303 nucleotides (cucumber strain), i.e. about 40-70 nucleotides fewer than the viroids related to potato spindle tuber viroid. The extent of sequence homology between hop stunt and potato spindle tuber viroids is 55% (Ohno et al., 1983).
Stability in Sap
In crude sap from diseased cucumber, the hop strain is not inactivated by heating at 84°C for 10 min or by storage for 3 days at 4°C (Sasaki & Shikata, 1978). The hop strain was not inactivated completely, even when razor blades contaminated with LiCl-soluble fraction from hop sap were heated for 10 min at 140°C and then used to cut plants (Takahashi & Yaguchi, 1985).
Purification
Blend frozen tissues of infected cucumber in 1.5 vol. 1 M K2HPO4, containing 1% SDS and 0.1% bentonite, and 0.2 vol. of a 1:1 mixture of phenol and chloroform. Collect the aqueous phase by centrifugation and precipitate the nucleic acid from 70% ethanol. Treat the nucleic acid preparation with an equal volume of 2-methoxyethanol, 2% cetyltrimethyl ammonium bromide, 2 M LiCl and 25 g/ml of DNase I. Apply the nucleic acid, in buffer containing 35% ethanol, to a column of CF-11 cellulose, wash the column with the same buffer containing 35% ethanol and then elute the viroid with ethanol-free buffer. Combine the resulting low M. Wt RNA fractions and purify by polyacrylamide gel electrophoresis (7.5, 10, 12.5 or 20% gels are effective) under non-denaturing conditions. Excise the band specific for hop stunt viroid from the gel and extract viroid RNA (Fig. 6) by maceration and elution of the gel. Further purification may be performed by polyacrylamide gel electrophoresis under denaturing conditions (7-8 M urea) if necessary. About 20-40 µg hop stunt viroid RNA is obtained from 1 kg infected cucumber leaves (Yoshikawa & Takahashi, 1982; Ohno et al., 1982; Uyeda et al., 1984).
Properties of Infective Nucleic Acid
Hop stunt viroid is more readily extracted in 0.5 M than in 0.005 M phosphate buffer (Sasaki & Shikata, 1978; Takahashi & Takusari, 1979). It is sensitive to RNase, but not to DNase (Sasaki & Shikata, 1978). Its sedimentation coefficient is 6-9 S, estimated by centrifugation in 5-20% sucrose density gradients (Sasaki & Shikata, 1978). When electrophoresed under denaturing conditions (Fig. 6), hop stunt viroid separates into circular (Fig. 7) and linear molecules; both forms are infective on cucumber (Yoshikawa & Takahashi, 1982; Ohno et al., 1982; Uyeda et al., 1984). Double-stranded cDNAs consisting of copies of 2 to 4 unit lengths of hop stunt viroid RNA are infective, and cause symptoms similar to those induced by native hop stunt viroid RNA (Meshi et al., 1984) (Fig. 4).
Molecular Structure
The molecule of the hop strain is a covalently closed single-stranded RNA of 297 nucleotides (Ohno et al., 1983). The complete nucleotide sequence is known (Ohno et al., 1983) (Fig. 8). The molecules of the grapevine (4 isolates) and cucumber strains consist of 297 and 303 nucleotides respectively, and differ from the hop strain by 1 and 16 nucleotide replacements, respectively (Sano et al., 1984, 1985, 1986) (Fig. 8).
Relations with Cells and Tissues
Studies on the subcellular distribution of the agent indicate that it is localized in the nucleus and plasma membrane (Takahashi et al., 1982). Electron microscopic examination shows that infection causes typical aberrations of the cell walls in all tissues of infected cucumber leaves (Momma & Takahashi, 1982; Kojima et al., 1983). Another type of alteration in infected leaf cells is the disintegration of the chloroplasts and marked changes of the membrane system (Kojima et al., 1983).
Notes
The herbaceous host range of the hop stunt viroid seems restricted to cucurbitaceous and solanaceous plants; no hosts in other families were detected in hop gardens in Japan (Yaguchi & Takahashi, 1984). However, grapevine and hop share the same cultivated area in Japan, and more than 80% of grapevine plants are infected with the grapevine strain (Sano et al., 1986). These facts suggest that the viroid in hop may have originated from the one in grapevine. Recently, a similar viroid was found in citrus (Citrus medica), plum (Prunus salicina) and peach (Prunus persica) in Japan (Sano et al., 1986a, 1986b). The potential distribution of hop stunt and related viroids seems to be world-wide.
Figures

Symptoms of hop stunt disease in the field. The main bines of affected plants (arrow) are severely shortened.

Cones of (left) healthy and (right) diseased hop. (From Yamamoto et al., 1973.)

Symptoms on cucumber flowers: (above), healthy; (below), infected with a hop strain.

Symptoms on cucumber plants (cv. Suyo) 6 weeks after inoculation: (left), healthy; (middle), inoculated with a hop strain; (right), inoculated with cloned cDNA of a hop strain.
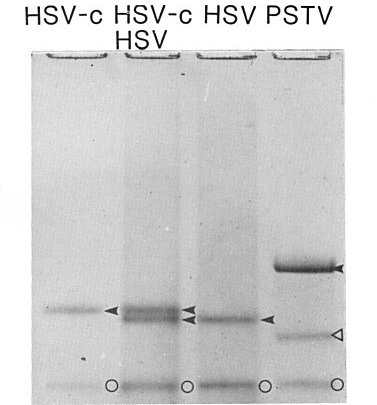
Electrophoresis of preparations of hop (HSV) and cucumber (HSV-c) strains of hop stunt viroid and of potato spindle tuber viroid (PSTV) on 5% polyacrylamide gel under denaturing conditions (7 M urea). The circular molecules of the hop strain migrate slightly faster than those of the cucumber strain in these conditions. The circular molecules of each viroid are indicated by black arrowheads; the linear molecules of PSTV are indicated by a white arrowhead and molecules of healthy plant RNA are indicated by circles (O). The linear molecules of HSV and HSV-c cannot be seen because of their low concentration (Sano et al., 1984).

Patterns of low M. Wt RNA species from healthy cucumber (left) and from cucumbers infected with a cucumber (HSV-c) strain (right), separated by electrophoresis in 15% polyacrylamide gel under non-denaturing conditions. One main specific band (arrow) is produced by the viroid-containing preparation (Uyeda et al., 1984).

Electron micrograph of circular molecules of the hop strain denatured in 90% formamide at 60°C. Bar represents 100 nm.

Secondary structures of the hop strain (top) (Ohno et al., 1983; sequence X00009), the grapevine strain (middle) (Sano et al., 1985; sequence E01844) and the cucumber strain (bottom) (Sano et al., 1984; sequence X00524). In the two lower diagrams, the nucleotides different from those of the hop strain are indicated by arrows.
References list for DPV: Hop stunt viroid (326)
- Kojima, Murai & Shikata, J. Fac. Agric. Hokkaido Univ. 61: 219, 1983.
- Meshi, Ishikawa, Ohno, Okada, Sano, Uyeda & Shikata, J. Biochem. 95: 1521, 1984.
- Momma & Takahashi, Phytopath. Z. 104: 211, 1982.
- Ohno, Akiya, Higuchi, Okada, Yoshikawa, Takahashi & Hashimoto,Virology 118: 54, 1982.
- Ohno, Takamatsu, Meshi & Okada, Nucleic Acids Res. 11: 6185, 1983.
- Sano, Sasaki & Shikata, Ann. phytopath. Soc. Japan 47: 599, 1981.
- Sano, Uyeda & Shikata, Ann. phytopath. Soc. Japan 50: 339, 1984.
- Sano, Uyeda, Shikata, Ohno & Okada, Nucleic Acids Res. 12: 3427, 1984.
- Sano, Ohshima, Uyeda, Shikata, Meshi & Okada, Proc. Japan Acad. 61B: 265, 1985.
- Sano, Ohshima, Hataya, Uyeda, Shikata, Chou, Meshi & Okada, J. gen.Virol. 67: 1673, 1986.
- Sano, Hataya, Sasaki & Shikata, Proc. Japan Acad. 62B: 325, 1986a.
- Sano, Hataya, Terai & Shikata, Proc. Japan Acad. 62B: 98, 1986b.
- Sano, Kudo, Sugimoto & Shikata, J. Virol. Meth. 19: 109, 1988.
- Sasaki & Shikata, Proc. Japan Acad. 53B: 103, 1977a.
- Sasaki & Shikata, Proc. Japan Acad. 53B: 109, 1977b.
- Sasaki & Shikata, Ann. phytopath. Soc. Japan 44: 405, 1978.
- Sasaki & Shikata, Rev. Pl. Prot. Res. 13: 97, 1980.
- Shikata, Sano & Uyeda, Proc. Japan Acad. 60B: 202, 1984.
- Takahashi & Takusari, Phytopath. Z. 96: 352, 1979.
- Takahashi & Yaguchi, Z. PflKrankh. PflSchutz. 92: 132,1985.
- Takahashi, Yaguchi, Oikawa & Kamita, Phytopath. Z. 103: 285, 1982.
- Uyeda, Sano & Shikata, Ann. phytopath. Soc. Japan 50: 331, 1984.
- Van Dorst & Peters, Neth. J. Pl. Path. 80: 85, 1974.
- Yaguchi & Takahashi, Phytopath. Z. 109: 32, 1984.
- Yamamoto, Kagami, Kurokawa, Nishimura, Kubo, Inoue & Murayama, Mem. Fac. Agric. Hokkaido Univ. 7: 491, 1970.
- Yamamoto, Kagami, Kurokawa, Nishimura, Ukawa & Kubo, Rep. Res. Lab. Kirin Brewery Co. Ltd. 16: 49, 1973.
- Yoshikawa & Takahashi, Ann. phytopath. Soc. Japan 48: 182, 1982.