Details of DPV and References
DPV NO: 338 September 1988
Family: Potyviridae
Genus: Potyvirus
Species: Iris severe mosaic virus | Acronym: ISMV
This is a revised version of DPV 147
Iris severe mosaic virus
A. A. Brunt Institute of Horticultural Research, Littlehampton, Sussex, England, BN17 6LP
A. F. L. M. Derks Bulb Research Centre, 2160 AB Lisse, The Netherlands
O. W. Barnett Clemson University, Clemson, South Carolina 29634-0377, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described in bulbous and rhizomatous irises by Brierley & McWhorter (1934, 1936) and in crocus by Van Slogteren (1956, 1958a). Virus partially characterized by Barnett et al. (1971), Brunt & Phillips (1980) and Derks et al. (1985).
Selected synonyms
- Bearded iris mosaic virus
(Brierley & Smith, 1948; Rev. appl. Mycol. 36: 763)
- Beardless iris mosaic virus (Brierley & Smith, 1948)
- Irisgrijsvirus (Van Slogteren, 1958a)
- Iris mosaic virus (Rev. appl. Mycol. 13: 380)
- Iris yellow mosaic virus (Rev. appl. Mycol. 43: 881e)
- Beardless iris mosaic virus (Brierley & Smith, 1948)
-
A virus with fiexuous filamentous particles c. 750 nm long, prevalent in bulbous and rhizomatous irises and occasionally present in crocus. It is transmissible in the non-persistent manner by aphids and, less easily, by mechanical inoculation. World-wide distribution.
Main Diseases
Causes conspicuous chlorotic stripes and/or mosaic patterns in the leaves, and breaking in the flowers of intolerant cultivars of bulbous iris (Fig. 1, Fig. 2), rhizomatous iris (bearded, crested and beardless types; Fig. 4, Fig. 5) and Crocus vernus (Fig. 3). Symptoms are usually more severe in plants grown at temperatures of 16°C or less, or in those also containing other viruses (Brunt, 1975; Barnett et al., 1971; Asjes, 1979; Derks et al., 1985).
Geographical Distribution
Probably present wherever bulbous and rhizomatous irises are grown; reported from many countries, including Australia (Carne, 1927), The Netherlands (Atanosoff, 1928), USA (Griffiths, 1928; Brierley & McWhorter, 1934), Japan (Fukushi, 1932), Sweden (Ingelström, 1938), Denmark (Neergaard, 1939), Czechoslovakia (Blattny, 1938), Israel (Loebenstein & Alper, 1963) and Britain (Ainsworth, 1937). Found also in I. anglica (= Xiphium latifolium) growing wild in the Spanish Pyrenees (A. F. L. M. Derks, unpublished information). Known to occur in crocus in the Netherlands (Van Slogteren, 1956) and Britain (A. A. Brunt & S. Phillips, unpublished information).
Host Range and Symptomatology
The virus is transmissible by aphids and by inoculation of sap to its three natural hosts and to Belamcanda chinensis, but to none of 30 species from 7 dicotyledonous families (Barnett et al., 1971; Brunt, 1975; Lisa, 1980; Derks et al., 1985).
- Diagnostic species
Belamcanda chinensis. Commonly-occurring isolates of the type strain from rhizomatous iris induce a few spreading necrotic lesions in inoculated leaves after c. 14 days and severe chlorotic mottling in the first few systemically infected leaves (Fig. 6), but symptomless infection in leaves produced later; avirulent isolates induce inconspicuous or symptomless systemic infection, but virulent isolates cause conspicuous leaf chlorosis (Fig. 7), severe stunting, leaf necrosis and death of infected plants (Brierley & Smith, 1948; Barnett et al., 1971; Travis, 1957; Derks et al., 1985; Derks & Hollinger, 1986).
Crocus vernus. Pale green mosaic patterns, particularly at leaf bases, and dark stripes in the petals (Fig. 3) in the year after inoculation.
Rhizomatous irises (Pogon, Apogon, Oncocyclus and Evansia types). Intolerant cultivars show conspicuous chlorotic mosaic on the leaves (Fig. 4), and produce short-lived flowers which are often broken or have ‘tear-drop’ markings (Fig. 5); some tolerant cultivars remain almost symptomless, others have faintly chlorotic leaves but normal flowers. The leaves of I. tectorum (crested iris) often develop chlorotic or water-soaked ringspot and line patterns.
Bulbous irises (especially Iris x hollandica). Pale green or yellow stripes at the leaf bases, extending upward from below soil level, and tear-drop markings or stripes in the petals (Fig. 1, Fig. 2).
- Propagation species
Cultures can be maintained conveniently in Belamcanda chinensis, Iris x hollandica and Crocus vernus. All three species are also good sources of inoculum and of virus for purification.
- Assay species
No species are sufficiently susceptible for sensitive assays. Iris x hollandica and Belamcanda chinensis, however, can be used for aphid transmission and virus stability tests.
Strains
Three strains of the virus have been recognized by the severity of the symptoms they induce in Belamcanda chinensis (Barnett & Brunt, 1975; Derks & Hollinger, 1986).
Type strain. In B. chinensis this strain induces a few local necrotic lesions followed initially by conspicuous systemic leaf chlorosis; leaves produced later are often symptomless. Common in bearded Eupogon irises (e.g. Iris pumila and I. ricardi), but also detected in I. gatesii (Oncocyclus iris), I. x hollandica and I. anglica.
Avirulent strain. In B. chinensis this strain induces distinct necrotic or chlorotic stripes and/or ringspots in inoculated leaves, but almost symptomless infection in systemically infected leaves. Isolated from I. x hollandica, and also with difficulty from I. susiana (bearded Oncocyclus iris) and Crocus vernus.
Virulent strain. In B. chinensis, isolates of this strain from I. spuria (beardless iris) induce severe leaf chlorosis (Fig. 6) and eventual death of systemically infected leaves. Symptoms in I. spuria and B. chinensis are indistinguishable from those previously attributed to ‘beardless iris mosaic virus’ (Brierley & Smith, 1948). Virulent strains have also been isolated from I. aurea, I. germanica and I. susiana (Fig. 7).
Transmission by Vectors
Transmitted efficiently by the aphids Macrosiphum euphorbiae and Myzus persicae after acquisition feeds of 3-5 min, but not by Aulacorthum solani or Aulacorthum (Neomyzus) circumflexum nor by the bulb aphids Dysaphis tulipae or Rhopalosiphoninus tulipaella (Brierley & McWhorter, 1934; Brierley & Smith, 1948; Travis, 1957; Barnett et al., 1971; Asjes, 1979). Spread of the virus in bulbous iris can be partially controlled by spraying crops with mineral oils (Deutsch & Loebenstein, 1967) or pyrethroid insecticide (Asjes, 1985).
Transmission through Seed
Not seed-borne in Belamcanda chinensis (Barnett et al., 1971).
Serology
The virus is a good immunogen: antisera with titres of 1/1280 to 1/16,000 in tube precipitin tests are readily obtained by injecting rabbits intramuscularly with virus emulsified with either Freund’s complete or Freund’s incomplete adjuvant (Derks et al., 1980; Barnett et al., 1971; A. A. Brunt, unpublished data.)
Relationships
The virus has many properties in common with potyviruses, but it is serologically unrelated
to similar viruses commonly infecting iridaceous species
(iris mild mosaic,
iris fulva mosaic,
bean yellow mosaic,
turnip mosaic,
freesia mosaic) or to four other potyviruses
(Barnett et al., 1971;
Brunt, 1975;
Derks & Hollinger, 1986).
Although iris severe mosaic virus isolates from bearded and bulbous irises are serologically related (Derks & Hollinger, 1986; O. W. Barnett, unpublished data; A. A. Brunt, unpublished data), their RNA molecules showed no sequence homology in reciprocal molecular hybridization tests with cDNA prepared to the nucleic acid of each (O. W. Barnett, unpublished data). The failure to detect hybridization might indicate that ‘bearded iris mosaic virus’ is not synonymous with iris severe mosaic virus; the hybridization data, however, need to be confirmed because serological and hybridization tests usually give similar information about the relationships between viruses, strains and isolates.
Stability in Sap
In Belamcanda chinensis sap, the thermal inactivation point is 50°C, dilution end-point 10-3, and infectivity is lost at 27°C after 24-36 h (Barnett et al., 1971).
Purification
Yields of over 5 mg/kg leaf tissue of mostly unaggregated virus are obtained as described
by Huttinga (1973).
Homogenize infected Belamcanda chinensis or Iris x
hollandica leaves (1 g/3 ml) in 0.1 M Tris thioglycollate buffer (pH 8.9) and 1.6 vol of
a 1:1 mixture of carbon tetrachloride and chloroform, subject the aqueous phase to one or two
cycles of differential centrifugation (1.5 h at 44,000 g; 10 min at 5000
g), and further purify the virus by rate zonal sucrose density gradient
centrifugation.
The virus can also be purified from Crocus vernus by clarifying leaf extracts with chloroform before precipitating virus with polyethylene glycol in a procedure similar to that used for purifying lily symptomless virus (Derks & Vink-van den Abeele, 1980); preparations so obtained are relatively free of host contaminants and are thus especially useful for the production of antisera for enzyme-linked immunosorbent assays (A. F. L. M. Derks, unpublished information).
Properties of Particles
Sedimentation coefficient (s°20, w): c. 150 S; some preparations may also contain faster sedimenting virus aggregates (A. A. Brunt, unpublished information).
A260/A280: 1.12; A260(max)/A280(min): 1.15 (both values after correction for light-scattering).
Particle Structure
Particles (Fig. 8) are slightly flexuous filaments mostly measuring c. 750 x 12 nm. A central canal and the helical arrangement of the protein subunits can be seen in some particles mounted in uranyl formate.
Particle Composition
Nucleic acid: RNA; c. 5% of particle weight (estimated spectrophotometrically).
Protein: c. 95% of particle weight; one type of polypeptide of M. Wt 33,000 (Alper et al., 1984).
Relations with Cells and Tissues
In ultrathin sections of infected iris leaves and petals, the virus particles are often scattered throughout the cytoplasm, but in Belamcanda chinensis cells they are also sometimes found in membrane-enclosed ovoid masses c. 800 nm in diameter (Fig. 9). Infected iris and Belamcanda chinensis cells also contain inclusions (Fig. 10) seen in sections as pinwheels, bundles and laminated aggregates (Barnett et al., 1971; Lisa, 1980). The inclusion protein is composed of a single polypeptide, of M. Wt 71,000 (Alper et al., 1984; Hammond & Lawson, 1985).
Notes
More than 50 years ago, Brierley & McWhorter (1934) showed that severe leaf chlorosis and flower-breaking, both in bulbous and in rhizomatous irises, were induced by one and the same virus. It was later suggested, however, that the virus (‘bearded iris mosaic virus’) commonly infecting rhizomatous irises was distinct from that (iris severe mosaic virus) prevalent in bulbous irises (Brierley & Smith, 1948; Travis, 1957). More recently it has been confirmed that iris severe mosaic virus induces conspicuous symptoms in both bulbous and rhizomatous irises (Derks & Hollinger, 1986). For historical and other reasons, the name iris severe mosaic virus has precedence over bearded iris mosaic virus and other synonyms (Hammond et al., 1985).
Four potyviruses are known to infect bulbous and rhizomatous irises (Brunt & Phillips, 1980; Derks et al., 1985; Inouye & Mitsuhata, 1978). Unlike iris severe mosaic virus, the other three (bean yellow mosaic, iris mild mosaic and turnip mosaic viruses) are all readily sap-transmitted to dicotyledonous diagnostic hosts; moreover, all three are serologically distinct from iris severe mosaic virus. The latter is also serologically distinct from another potyvirus (iris fulva mosaic virus) infecting rhizomatous irises (Barnett & Alper, 1977). Two other viruses with filamentous particles, narcissus latent carlavirus (Brunt, 1977; Asjes, 1979) and lilac chlorotic leafspot closterovirus (Lisa, 1980), also infect irises but have particles which differ from those of iris severe mosaic virus in length, in serological reactions and in their ability to infect dicotyledonous hosts.
Rhizomatous irises infected with iris severe mosaic virus in Britain also commonly contain
cucumber mosaic cucumovirus
(Brunt, 1968) or, rarely,
broad bean wilt fabavirus
(Bailiss et al., 1975).
Tobacco ringspot nepovirus occurs occasionally in rhizomatous iris in the
USA and Britain
(Travis & Brierley, 1957;
Brunt, 1972) and in bulbous iris in The
Netherlands
(Asjes, 1979).
Tobacco mosaic tobamovirus and
tobacco rattle tobravirus have also
been reported to occur in bulbous iris in The Netherlands
(Asjes, 1979). Unlike iris severe
mosaic virus, however, these five viruses have isometric or rod-shaped particles, they infect
a wide range of plant species and all can be quickly identified by their serological or other
properties.
Iris severe mosaic virus occurs naturally in Crocus vernus and C. flavus; bean yellow mosaic virus, however, occurs in C. flavus and C. sativus but not in C. vernus (Derks et al., 1983; Russo et al., 1979). Bean yellow mosaic and other viruses reported to infect crocus such as tobacco rattle (Van Slogteren, 1958b), arabis mosaic (Asjes, 1974) and cucumber mosaic (A. A. Brunt & S. Phillips, unpublished information) are easily distinguished from iris severe mosaic virus in having extensive natural and experimental host ranges and, with the exception of the serologically distinct bean yellow mosaic virus, in having isometric or rod-shaped particles.
Figures

Conspicuous systemic leaf chlorosis in bulbous iris cv. Professor Blaauw.

Flower breaking in bulbous iris cv. Professor Blaauw.

Flower breaking in Crocus vernus cv. Remembrance.

Mosaic leaf symptoms in bearded iris.

‘Tear drop’ markings in infected flower of bearded iris.

Severe systemic leaf chlorosis in Belamcanda chinensis induced by virulent strains from Iris spuria.

Severe systemic leaf chlorosis in Belamcanda chinensis induced by virulent strains from Iris susiana.

Virus particles mounted in potassium phosphotungstate. Bar represents 250 nm.
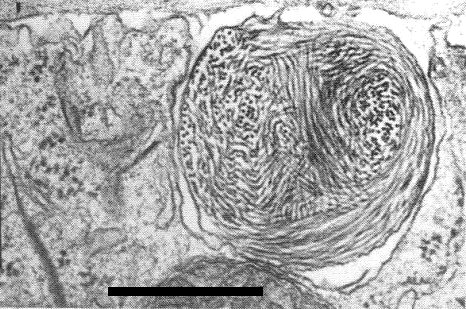
Membrane-bound aggregate of virus particles. Bar represents 500 nm.

Section of Belamcanda chinensis leaf showing pinwheels and laminated aggregates. Bar represents 500 nm.
References list for DPV: Iris severe mosaic virus (338)
- Ainsworth, Rep. exp. Res. Stn, Cheshunt for 1936: 62, 1937.
- Alper, Salomon & Loebenstein, Phytopathology 74: 960, 1984.
- Asjes, Agric. Environm. 1: 303, 1974.
- Asjes, Neth. J. Pl. Path. 85: 269, 1979.
- Asjes, Crop. Prot. 4: 485, 1985.
- Atanasoff, Izv. búlg. bot. Druzh. 2: 51, 1928.
- Bailiss, Brunt & Dale, Pl. Path. 24: 60, 1975.
- Barnett & Alper, Phytopathology 67: 448, 1977.
- Barnett & Brunt, CMI/AAB Descr. Pl. Viruses 147, 4 pp., 1975.
- Barnett, de Zoeten & Gaard, Phytopathology 61: 926, 1971.
- Blattny, Ochr. Rost. 14: 86, 1938.
- Brierley & McWhorter, Phytopathology 24: 4, 1934.
- Brierley & McWhorter, J. agric. Res. 53: 621, 1936.
- Brierley & Smith, Phytopathology 38: 574, 1948.
- Brunt, Rep. Glasshouse Crops Res. Inst., 1967: 101, 1968.
- Brunt, Rep. Glasshouse Crops Res. Inst., 1971: 119, 1972.
- Brunt, Acta Hort. 47: 45, 1975.
- Brunt, Ann. appl. Biol. 87: 335, 1977.
- Brunt & Phillips, Acta Hort. 109: 503, 1980.
- Carne, J. Proc. R. Soc. West. Aust. 14: 23, 1927.
- Derks & Hollinger, Acta. Hort. 177: 555, 1986.
- Derks & Vink-van den Abeele, Neth. J. Pl. Path. 86: 239, 1980.
- Derks, Vink-van den Abeele & Muller, Acta Hort. 110: 31, 1980.
- Derks, Vink-van den Abeele & Hollinger, Jversl. Lab. Bloembollenonderz., Lisse, 1982: 61, 1983.
- Derks, Hollinger & Vink-van den Abeele, Acta Hort. 164: 309, 1985.
- Deutsch & Loebenstein, Pl. Dis. Reptr 51: 318, 1967.
- Fukushi, Trans. Sapporo Nat. Hist. Soc. 12: 130, 1932.
- Griffiths, Circ. U.S. Dep. Agric. 25: 16, 1928.
- Hammond & Lawson, Acta Hort. 164: 225, 1985.
- Hammond, Brunt, Derks, Inouye, Barnett, Allen & Lawson, Acta Hort. 164: 395, 1985.
- Huttinga, Neth. J. Pl. Path. 79: 125, 1973.
- Ingelström, Växtskyddnotiser 2: 22, 1938.
- Inouye & Mitsuhata, Nogaku Kenkyu 57: 1, 1978.
- Lisa, Acta Hort. 110: 39, 1980.
- Loebenstein & Alper, Phytopathology 53: 349, 1963.
- Neergaard, Arsberetn. J. E. Ohlsens Enkes plpatol. Lab., 1938-1939, 16 pp., 1939.
- Russo, Martelli, Cresti & Ciampolini, Phytopath. Mediterranea 18: 189, 1979.
- Travis, Phytopathology 47: 454, 1957.
- Travis & Brierley, Pl. Dis. Reptr 41: 524, 1957.
- Van Slogteren, Jversl. Lab. Bloembollenonderz., Lisse, 1955: 82, 1956.
- Van Slogteren, Jversl. Lab. Bloembollenonderz., Lisse, 1957: 66, 1958a.
- Van Slogteren, Tijdschr. PlZiekt 64: 452, 1958b.