Details of DPV and References
DPV NO: 340 December 1989
Family: Potyviridae
Genus: Potyvirus
Species: Johnsongrass mosaic virus | Acronym: JGMV
Johnsongrass mosaic virus
D. D. Shukla CSIRO, Division of Biotechnology, Parkville Laboratory, 343 Royal Parade, Parkville, Victoria 3052, Australia
D. S. Teakle Department of Microbiology, University of Queensland, St. Lucia, Qld. 4067, Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Originally described by Taylor & Pares (1968) as Australian maize dwarf mosaic virus and by Teakle & Grylls (1973) as Australian Johnsongrass strain of sugarcane mosaic virus, but later shown (Shukla et al., 1987) to be an independent member of the potyvirus group and named Johnsongrass mosaic virus.
Synonyms
- Australian Johnsongrass strain of sugarcane mosaic virus
(Teakle & Grylls, 1973)
- Australian maize dwarf mosaic virus (Taylor & Pares, 1968)
- U.S. oat strain of maize dwarf mosaic virus (McDaniel & Gordon, 1985)
- Australian maize dwarf mosaic virus (Taylor & Pares, 1968)
-
A virus with flexuous filamentous particles c. 750 nm long, containing single-stranded RNA. It is transmissible mechanically, and by several aphid species in a non-persistent manner, and induces the formation of cytoplasmic cylindrical (pinwheel and bundle) inclusions in host cells. It is known to occur in mainland Australia and in USA, where it causes mosaic and necrotic diseases of sorghums and mosaic diseases of maize and various grasses.
Main Diseases
Grain, fodder and weed sorghums (Sorghum almum, S. bicolor and S. sudanense), and maize and sweet corn (Zea mays), are important natural hosts, developing mosaic or necrotic foliage symptoms. Stunting and yield reduction in sorghum are greater in plants that react with necrosis than in those that react with mosaic (Teakle et al., 1970; Henzell et al., 1979). Some maize and sweet corn lines and hybrids become chlorotic and stunted (Grogan & Teakle, 1969; Persley et al., 1981). Resistance to the type strain is available in Krish sorghum (Teakle & Pritchard, 1971; Condé et al., 1976; Henzell et al., 1982; Persley et al., 1985) or Q7539 sorghum (Persley et al., 1972; Henzell et al., 1982) and in several maize lines (Grogan & Teakle, 1969; Persley et al., 1981). However, a strain reported from Queensland can infect sorghums carrying Krish-type resistance (D. M. Persley, R. S. Greber & R. G. Henzell, unpublished data).
Geographical Distribution
Limited. Reported from Australia and USA. In Australia it is widespread in New South Wales and Queensland, and of restricted distribution in Victoria and the Northern Territory (Teakle & Grylls, 1973; Penrose, 1974a). In USA it has been found in Texas (McDaniel & Gordon, 1985; Shukla et al., 1989b).
Host Range and Symptomatology
The main weed hosts in Queensland and New South Wales are Johnsongrass (Sorghum halepense) and a wild sorghum (S. verticilliflorum), which develop mosaic and ringspot symptoms and act as perennial reservoirs of the virus (Taylor & Pares, 1968; Teakle & Grylls, 1973; Penrose, 1974a). Commercial fodder and grain sorghums, such as S. almum, S. bicolor and S. sudanense, develop mosaic (Fig. 1), ringspot or necrosis (Fig. 2) symptoms, and sometimes stunting. Whether the symptoms are mosaic or necrosis is genetically controlled (Teakle et al., 1970; Penrose, 1974b; Persley et al., 1977). The ‘red stripe’ necrotic reaction depends on the N gene and occurs at both high and low temperatures, whereas the ‘red leaf’ necrotic reaction depends on the rlf gene and needs low temperature (e.g. 15°C) for expression (Persley et al., 1977). Maize and sweet corn develop mosaic, ringspotting (Fig. 3) and chlorosis. Other natural graminaceous hosts include Brachiaria miliiformis, B. praetervisa, Cenchrus ciliaris, Dinebra retroflexa, Echinochloa colona, E. crus-galli, E. frumentacea, Eragrostis cilianensis, Panicum miliaceum, Paspalum orbiculare, Pennisetum typhoideum, Setaria anceps, S. italica and S. verticillata (Teakle & Grylls, 1973; Penrose, 1974a; Persley & Greber, 1977). Many other grasses have been shown to be susceptible following mechanical inoculation, but some do not develop symptoms (Penrose, 1974a); no monocotyledonous hosts outside the Gramineae, nor any dicotyledonous hosts, have been found (Teakle & Grylls, 1973).
-
Diagnostic species
- Sorghum bicolor
OK Y8 and other lines with the N gene in the dominant state develop necrotic red stripe symptoms followed by a severe yellow mosaic and stunting (Fig. 2; Teakle et al., 1970). This reaction allows the virus to be distinguished from sugarcane mosaic, maize dwarf mosaic and sorghum mosaic viruses (see Notes).Propagation species
- Sorghum bicolor
lines which have the nn genetic constitution and react with mosaic symptoms are suitable, as are susceptible lines of Zea mays such as Iochief sweet corn.Assay species
- Although Sorghum bicolor lines with the N gene in the dominant state give necrotic local lesions on inoculation (Teakle et al., 1970), these local lesions have not been used in assays. Young S. bicolor or Zea mays seedlings can be examined for systemic symptoms following inoculation.
Strains
Type strain. Endemic in Australia in Johnsongrass, maize and sorghum
(Teakle & GryIls, 1973;
Shukla et al., 1987).
Krish-infecting strain. Differs from the type strain in being able to infect sorghum
with the Krish resistance gene in the dominant state (D. M. Persley, R. S. Greber & R. G.
Henzell, unpublished data). No serological differences have been observed between this and the
type strain (D. D. Shukla, unpublished data).
Texas strain. Isolated only once from Texas, USA, and originally referred to as maize dwarf mosaic virus-O by McDaniel & Gordon (1985) because unlike other strains of aphid-borne potyviruses infecting species of Gramineae in the USA, it was able to infect oats. However, the type strain of Johnsongrass mosaic virus has also been found recently to infect oat cultivars (Tosic et al., 1990). No differences were observed in host response when the type and the Texas strains of Johnsongrass mosaic virus were compared on a set of eleven sorghum inbred lines (Tosic et al., 1990). The two strains are also very closely related serologically but can be distinguished by using monoclonal antibodies (D. D. Shukla & D. R. Hewish, unpublished data).
Transmission by Vectors
The four aphid species shown to transmit are Aphis craccivora, Aphis gossypii, Myzus persicae and Rhopalosiphum maidis. In these tests the aphids were given either acquisition feeding times of 24 h and inoculation feeding times of 24-48 h or a single probe for acquisition and 1 h for inoculation (Teakle & Grylls, 1973). The latter type of test suggests that the virus-vector relations are of the non-persistent type.
Transmission through Seed
This was not detected in 500 sweet corn seedlings grown from seed collected from infected parents (Teakle & Grylls, 1973).
Serology
Antisera with homologous titres of 1/64-1/512 have been produced by injecting rabbits with purified preparations of particles. These antisera give flocculent precipitates when mixed with purified virus preparations in the microprecipitin test (Taylor & Pares, 1968; Teakle & Grylls, 1973; Penrose, 1974b; Shukla & Gough, 1979). Antisera can also be used in trapping, coating and gold labelling of particles for electron microscopic observations (Shukla & Gough, 1984; S. Srisink & D. S. Teakle, unpublished data). Antisera have also been produced to trypsin-resistant core particles and trypsin-resistant core protein (Shukla et al., 1988). A number of monoclonal antibody clones have been produced in mice by using intact particles as antigen (D. R. Hewish & D. D. Shukla, unpublished data).
Relationships
Johnsongrass mosaic virus is placed in the potyvirus group because of its filamentous particles c. 750 x 12 nm, its induction of pinwheel inclusions in cells, and its mechanical and aphid transmissibility. On the basis of an apparently distant serological relationship in microprecipitin tests, Johnsongrass mosaic virus was considered to be a strain of sugarcane mosaic virus (Taylor & Pares, 1968; Teakle & Grylls, 1973). However, more recent tests (microprecipitin, immunosorbent electron microscopy, electroblot immunoassay and enzyme-linked immunosorbent assay) have failed to confirm any close relationship (Penrose, 1974b; Shukla et al., 1983; Shukla & Gough, 1984; Hewish et al., 1986), and these two viruses are now considered distinct potyviruses (Shukla et al., 1989b).
Johnsongrass mosaic virus is serologically closely related to watermelon mosaic virus 2 and distantly related to bean yellow mosaic, maize dwarf mosaic, sorghum mosaic, sugarcane mosaic and clover yellow vein viruses (Shukla et al., 1988; 1989a; 1989b). Epitopes responsible for the relationship to watermelon mosaic virus 2 are contained in the surface-exposed N-terminal peptide region of the coat protein (Shukla et al., 1988) whereas epitopes related to those of bean yellow mosaic, maize dwarf mosaic, sorghum mosaic, sugarcane mosaic and clover yellow vein viruses are located in the trypsin-resistant core region of the coat protein. Indeed an antiserum to the trypsin resistant core region was found to react with every potyvirus tested (Shukla et al., 1988; 1989a). Monoclonal antibodies directed to the surface-exposed N-terminal region of the coat protein were found to be specific for Johnsongrass mosaic virus and its strains, whereas those directed to the trypsin-resistant core protein region possessed specificities for different biologically unrelated potyviruses. Some monoclonal antibodies reacted with all the potyviruses tested (D. R. Hewish & D. D. Shukla, unpublished data).
Stability in Sap
In slightly diluted sweet corn sap the thermal inactivation point is about 60°C, and the longevity in vitro at about 20°C is 2 days. The dilution end-point is about 10-4 (Teakle & Grylls, 1973).
Purification
Johnsongrass mosaic virus is reported to be easier to purify than sugarcane mosaic virus. However, aggregation of virus particles (Teakle & Pares, 1977; Fig. 4) is a problem.
1. (Taylor & Pares, 1968). This method is similar to that of Shepherd (1965) for sugarcane mosaic virus strains. Homogenize infected tissues with 0.5 M sodium citrate containing 0.5% mercaptoethanol (1 g tissue/1 ml). Clarify by emulsifying with chloroform (1:1) followed by low speed centrifugation. Filter through glass wool and pellet by ultracentrifuging 90 min at 30,000 rev./min. Repeat centrifugation cycle. Resuspend pellets in either 5 mM borate, pH 8.2 or, if emulsifying in Freund’s adjuvant, 50 mM phosphate buffer, pH 7·6. Penrose (1974b) used this method with the modification that only one-quarter volume of chloroform was used to emulsify the suspension.
2. (Teakle & Grylls, 1973). This method is similar to that of Snazelle et al. (1971) for sugarcane mosaic virus strains. Homogenize infected tissues in 17 mM ascorbic acid containing 10 mM NaDIECA (1 g tissue/2 ml) and strain through cheesecloth. Emulsify with chloroform (3 g tissue/1 ml) and centrifuge at low speed. Pellet by ultracentrifuging 2 h at 28,000 rev./min and resuspend pellets in 0.1 M borate buffer, pH 8.2. Clarify by low speed centrifugation and further purify by sucrose density gradient centrifugation.
3. (Gough & Shukla, 1981). Homogenize infected sweetcorn leaves in 1.5 vol. 0.5 M sodium borate buffer, pH 8.0, containing 0.15% (v/v) thioglycollic acid and 10 mM EDTA, and 0.5 vol. each of chloroform and carbon tetrachloride. Centrifuge the homogenate at 8000 g for 10 min, filter the supernatant fluid, add Triton X-100 to 5% (v/v) and stir for 30 min at 4°C. Centrifuge the mixture at 8000 g for 10 min, subject the supernatant fluid to two cycles of differential centrifugation (high speed - 100,000 g for 90 min, low speed - 8000 g for 10 min) and resuspend the pellets in 50 mM sodium borate buffer, pH 8.0, containing 10 mM EDTA. Purify further by centrifugation in a linear sucrose gradient (10-40%) prepared in the resuspension buffer. Unaggregated virus particles form a distinct band but a pellet of aggregated particles usually also forms at the bottom of the gradient. However, protein preparations made from either the band or the pellet each form a single band in SDS-PAGE, indicating that both fractions are devoid of host proteins. This procedure yields up to 100 mg virus/kg infected tissue.
Particle Structure
Particles are flexuous filaments c. 750 nm long and 12 nm wide (Fig. 5; Taylor & Pares, 1968; Teakle & Grylls, 1973).
Particle Composition
Nucleic acid: A single ssRNA species of c. 3.1 x 106 daltons (c. 9500 base pairs), c. 5% by particle weight (Gough, 1987).
Protein: A single polypeptide species of 33,510 daltons consisting of 303 amino acid
residues. The N-terminus is acetylated
(Shukla et al., 1987); 67 amino acid residues from
the N-terminus and 18 from the C-terminus of the coat protein are exposed on the surface of the
virus particle. Removal of these exposed regions does not affect the morphology or destroy
infectivity of the virus particles (Shukla et al., 1988).
Amino acid composition
(Shukla et al., 1987): ala 29, arg 17, asx 46, cys 2, glx 28,
gly 22, his 7, ile 12, leu 15, lys 21, met 12, phe 8, pro 14, ser 22, thr 22, trp 3, tyr 10 and
val 13.
The amino acid sequence is given by Gough et al. (1987) and Shukla et al. (1987).
Genome Properties
(Gough, 1987; Gough et al., 1987). The segment of the genome encoding the viral coat protein is adjacent to the 3' non-coding region. The nuclear inclusion protein gene is located upstream from the coat protein gene. The 3' non-coding region is 475 nucleotides long and is polyadenylated. The poly(A) tail consists of 50 adenosine residues. The RNA is translated in vitro into a number of protein products ranging in M. Wt from 25,000 to 92,000 daltons.
Relations with Cells and Tissues
The virus becomes systemic in many of its hosts. Virus particles and pinwheel and bundle inclusions are found in the cytoplasm of cells in infected tissues (Fig. 6). In sorghum they are equally common in chlorotic and green areas of infected leaves, but in sweet corn they are largely confined to the chlorotic areas. They are most numerous in mesophyll cells, but are also present in epidermal cells, sieve tubes, vascular parenchyma and companion cells. They are absent from chloroplasts, nuclei, mitochondria and cells of the chlorophyllous sheath surrounding the vascular bundle (Penrose, 1974c). Although nuclear inclusions, found in plants infected with some other potyviruses (e.g. tobacco etch and bean yellow mosaic viruses), have not been observed with Johnsongrass mosaic virus (Penrose, 1974c), presence of the nuclear inclusion gene in the Johnsongrass mosaic virus genome (Gough et al., 1987) suggests that nuclear inclusion protein is produced in infected tissue. Presumably the protein does not aggregate to form nuclear inclusion bodies as happens with some other potyviruses.
Notes
Recent studies on the taxonomy of aphid-borne potyviruses infecting species of Gramineae (Shukla et al., 1989b) have shown that virus isolates previously included as strains of sugarcane mosaic virus in fact comprise four distinct potyviruses. One of these is Johnsongrass mosaic virus (formerly known as Australian sugarcane mosaic virus strain JG and U.S. maize dwarf mosaic virus strain O); the other three are maize dwarf mosaic virus (U.S. strains A, D, E and F), sugarcane mosaic virus (U.S. strains A, B, D and E, Australian strains SC, BC and Sabi, and U.S. maize dwarf mosaic virus strain B) and sorghum mosaic virus (U.S. sugarcane mosaic virus strains H, I and M). The four viruses induce similar symptoms in some hosts, have host ranges usually restricted to the Gramineae and have some aphid vectors in common. However, they are serologically unrelated or only distantly related (Teakle & Grylls, 1973; Penrose, 1974c; Shukla & Gough, 1984; McDaniel & Gordon, 1985; Giorda et al., 1986; Hewish et al., 1986; Shukla et al., 1983; 1988; 1989a; 1989b). Previous reports of serological relationships among these viruses and their strains are caused by the presence in polyclonal antisera of antibodies to epitopes located in the conserved regions of potyvirus coat proteins; these antibodies recognize most potyviruses. The four viruses also differ considerably in the amino acid composition of their coat proteins (Hill et al., 1973; Von Baumgarten & Ford, 1981; Gough & Shukla, 1981; Shukla et al., 1987). The four viruses can also be distinguished by their reactions on certain sorghum inbred lines (Teakle & Grylls, 1973; Persley et al., 1985; Giorda et al., 1986; Tosic et al., 1990). Johnsongrass mosaic virus induces necrotic red stripes in sorghum lines OK Y8 and SA 8735 whereas the other three viruses induce only mosaic symptoms. Maize dwarf mosaic virus induces mosaic and necrotic spots in systemically infected leaves of cv. Rio (Exotic) sorghum, whereas the other three viruses induce either mosaic or symptomless infection. Strains of sugarcane mosaic virus induce necrotic stripes in inoculated leaves and mosaic and necrosis of new leaves of Atlas sorghum, whereas sorghum mosaic virus induces typical red leaf symptoms and the other two viruses induce only mosaic symptoms.
Acknowledgements
Figs 1 and 3 courtesy of Queensland Department of Primary Industries; Fig. 6 courtesy Sunee Srisink.
Figures

Part of a leaf of Sorghum bicolor showing mosaic symptoms.

Part of a leaf of S. bicolor cv. Yates NK220Y showing necrotic red stripe symptoms.
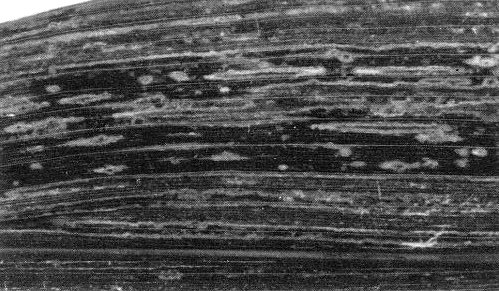
Part of a leaf of Zea mays showing mosaic and ringspotting.

Partially purified preparation negatively-stained with potassium phosphotungstate, with many particles aggregated. Bar represents 500 nm.

Particles extracted from a leaf of Zea mays and stained with potassium phosphotungstate. Bar represents 250 nm.
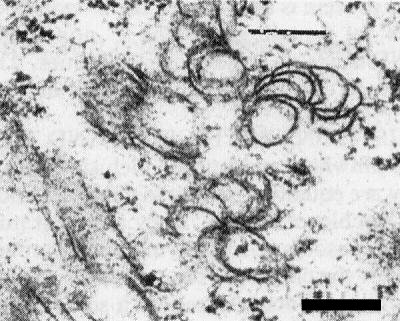
Pin-wheel lamellae in a section of a leaf of Zea mays. Bar represents 300 nm.
References list for DPV: Johnsongrass mosaic virus (340)
- Condé, Moore, Fletcher & Teakle, Aust. J. agric. Res. 27: 45, 1976.
- Giorda, Toler & Miller, Pl. Dis. 70: 624, 1986.
- Gough, Ph.D. thesis, Deakin Univ., Geelong, Australia, 116 pp., 1987.
- Gough & Shukla, Virology 111: 455, 1981.
- Gough, Azad, Hanna & Shukla, J. gen. Virol. 68: 297, 1987.
- Grogan & Teakle, Aust. J. exp. Agric. Anim. Husb. 9: 541, 1969.
- Henzell, Persley, Fletcher, Greber & van Slobbe, Aust. J. exp. Agric. Anim. Husb. 19: 225, 1979.
- Henzell, Persley, Greber, Fletcher & van Slobbe, Pl. Dis. 66: 900, 1982.
- Hewish, Shukla & Gough, J. virol. Methods 13: 79, 1986.
- Hill, Ford & Benner, J. gen. Virol. 20: 327, 1973.
- McDaniel & Gordon, Pl. Dis. 69: 602, 1985.
- Penrose, Aust. J. agric. Res. 25: 99, 1974a.
- Penrose, Pl. Dis. Reptr 58: 832, 1974b.
- Penrose, Phytopath. Z. 80: 157, 1974c.
- Persley & Greber, Aust. Pl. Path. Soc. Newsl. 6: 54, 1977.
- Persley, Greber & Moore, Aust. Pl. Path. Soc. Newsl. 1: 11-12, 1972.
- Persley, Moore & Fletcher, Aust. J. agric. Res. 28: 853, 1977.
- Persley, Martin & Greber, Aust. J. agric. Res. 32: 741, 1981.
- Persley, Henzell, Greber, Teakle & Toler, Pl. Dis. 69: 1046, 1985.
- Shepherd, Phytopathology 55: 1250, 1965.
- Shukla & Gough, J. gen. Virol. 45: 533, 1979.
- Shukla & Gough, Pl. Dis. 68: 204, 1984.
- Shukla, O'Donnell & Gough, Acta phytopath. Acad. Sci. hung. 18: 79, 1983.
- Shukla, Gough & Ward, Arch. Virol. 96: 59, 1987.
- Shukla, Strike, Tracy, Gough & Ward, J. gen. Virol. 69: 1497, 1988.
- Shukla, Jilka, Tosic & Ford, J. gen. virol. 70: 13, 1989a.
- Shukla, Tosic, Jilka, Ford, Toler & Langham, Phytopathology 79: 223, 1989b.
- Snazelle, Bancroft & Ullstrup, Phytopathology 61: 1059, 1971.
- Taylor & Pares, Aust. J. agric. Res. 19: 767, 1968.
- Teakle & Pares, in The Atlas of Insect and Plant Viruses, p. 317, ed.K. Maramorosch, New York: Academic Press, 478 pp., 1977.
- Teakle & Pritchard, Pl. Dis. Reptr 55: 596, 1971.
- Teakle & Grylls, Aust. J. agric. Res. 24: 465, 1973.
- Teakle, Moore, George & Byth, Aust. J. agric. Res. 21: 549, 1970.
- Tosic, Ford, Shukla & Jilka, Pl. Dis. 74: 549, 1990.
- Von Baumgarten & Ford, Phytopathology 71: 36, 1981.