Details of DPV and References
DPV NO: 348 December 1989
Family: Geminiviridae
Genus: Mastrevirus
Species: Miscanthus streak virus | Acronym: MiSV
Miscanthus streak virus
S. Yamashita Laboratory of Plant Pathology, Faculty of Agriculture, The University of Tokyo, Bunkyo-ku, Tokyo 113, Japan
Y. Doi Laboratory of Plant Pathology, Faculty of Agriculture, The University of Tokyo, Bunkyo-ku, Tokyo 113, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Yamashita et al. (1979, 1985).
- A virus with geminate particles c. 30 x 18 nm containing one species of circular single-stranded DNA. Reported only from the wild grass, Miscanthus sacchariflorus. No vector known. Not transmissible by mechanical inoculation
Main Diseases
The wild plant Miscanthus sacchariflorus (Gramineae) is the only known host. The virus causes leaf mosaic or streaking, sometimes ‘ragged leaf’ symptoms and stunting (Fig. 1), but is of no economic importance (Yamashita et al., 1985).
- A virus with geminate particles c. 30 x 18 nm containing one species of circular single-stranded DNA. Reported only from the wild grass, Miscanthus sacchariflorus. No vector known. Not transmissible by mechanical inoculation
Geographical Distribution
Reported only from local areas in Japan.
Host Range and Symptomatology
The virus has not been transmitted mechanically and its vector is unknown. Consequently it has not been transmitted to any species other than its sole natural host, M. sacchariflorus.
- Diagnostic and assay species
- None known.
- Propagation species
- Infected M. sacchariflorus is suitable for maintenance by vegetative propagation (rooted cuttings and subdivision of rootstocks) and is also a source of virus for purification.
Strains
No information.
Transmission by Vectors
No vector is known. The vector is probably a leafhopper, as with other geminiviruses occurring in Gramineae.
Transmission through Seed
No information.
Serology
The virus is moderately immunogenic. Antiserum was prepared in a rabbit given a course of one intramuscular and four intravenous injections of purified virus preparations. The antiserum had a titre of 1/128 in Ouchterlony double diffusion tests and gave a single precipitin line (Fig. 3). Immunosorbent electron microscopy has been used for virus detection (Yamashita et al., 1985).
Relationships
The virus is a typical geminivirus in its particle morphology and
physicochemical properties. In gel diffusion tests (Yamashita
et al., 1985; Fig. 3), no serological relationships were found to two
leafhopper-transmitted (subgroup A) geminiviruses, maize streak virus (Bock,
1974) and chloris striate mosaic virus (Francki & Hatta, 1980), or to two
whitefly-transmitted (subgroup B) geminiviruses, African cassava mosaic virus
(cassava latent virus) (Bock & Harrison, 1985) and tobacco leaf curl virus
(Osaki & Inouye, 1981).
The virus DNA has extensive sequence homology with those of maize streak virus and wheat dwarf virus.
Stability in Sap
Not known. As determined by electron microscopy, virus particle morphology was retained in leaf tissue stored at -80°C for at least 4 years (Yamashita et al., unpublished data).
Purification
(Yamashita et al., 1985). Store infected M. sacchariflorus leaves at -80°C. Homogenize in 4 vol. 0.1 M phosphate buffer (pH 5 .6) containing 0.1% thioglycollic acid and 1% Triton X-100. Express the homogenate through cheesecloth, clarify by treatment with 20% chloroform, and then subject to three cycles of differential centrifugation. Purify further by centrifugation in 10-40% sucrose density gradients.
Properties of Particles
Purified preparations showed a single light-scattering band in sucrose density gradient centrifugation. Sedimentation coefficient (s20,w): about 75 S (Yamashita et al., 1985; Fig. 2).
A260/A280: 1.6 (Yamashita et al., 1985).
Buoyant density in CsCl: 1.35 g/cm3 (Nonaka et al., 1985).
Particle Structure
Particles are geminate structures c. 30 x 18 nm (Yamashita et al., 1985), probably consisting of two incomplete T= 1 icosahedra with a total of 22 capsomeres (Fig. 4).
Particle Composition
Nucleic acid: One species of circular single-stranded DNA, M. Wt c. 0.87 x 106 (Nonaka et al., 1985; Ikeda et al., 1988; Chatani et al., 1988). The molecule has 2672 nucleotides, estimated by sequencing from cloned double-stranded DNA (Chatani et al., 1988). Buoyant density in CsCl is c. 1.72 g/cm3 (Nonaka et al., 1985).
Protein: A single polypeptide of M. Wt c. 28,000, estimated by SDS/polyacrylamide gel electrophoresis (Nonaka et al., 1985).
Genome Properties
The nucleotide sequence of the viral DNA has been determined (Chatani
et al., 1988); a hairpin-loop structure and AT-rich inverted repeat
were observed as in other geminiviruses.
Two potential open reading frames (ORF) starting with an ATG triplet and coding for polypeptides of more than 10 kilodaltons occur in the virus DNA (plus) strand (R) and two more in its complementary (minus) strand (L) (Fig. 8). There is an extensive amino acid sequence homology in the proteins coded by ORF RI and ORF Ll and the corresponding proteins of maize streak and wheat dwarf viruses (Chatani et al., 1988).
Relations with Cells and Tissues
In infected M. sacchariflorus, virus particles frequently occur in aggregates in the nucleus, also sometimes in the cytoplasm and vacuole. Their geminate nature is often obvious in the nucleus and vacuole. In the vacuole, unique geometrical crystalline arrays formed from geminate particles were also recognized. The geminate particles found in ultrathin sections were considered to be the natural form of the virus (Yamashita et al., 1985: Fig. 5, Fig. 6, Fig. 7).
Notes
Miscanthus streak virus is the only geminivirus reported from graminaceous plants in Japan. The virus has similar particle and genome properties to other leafhopper-transmitted (subgroup A) geminiviruses, but is serologically distinguishable from those tested. However, its vector is unknown. The virus can be distinguished from a potyvirus, sugarcane mosaic virus, known to occur in M. sacchariflorus by symptoms, particle morphology and lack of mechanical transmissibility (Yamashita et al., unpublished data).
Figures

Streaked (a) and ragged (b) leaf symptoms on naturally infected Miscanthus sacchariflorus.
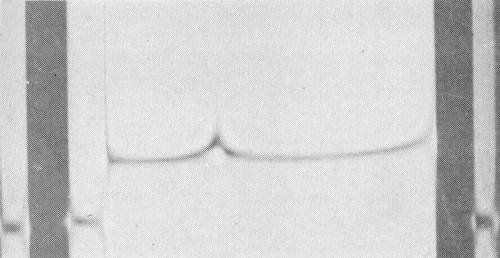
Schlieren pattern of purified preparation of the virus particles undergoing analytical ultracentrifugation. Sedimentation is from left to right.

Agar gel diffusion serological test. Antigen (Mi-V) is purified miscanthus streak virus. Antisera are: Mi-A, miscanthus streak virus; M-A, maize streak virus; C-A, chloris striate mosaic virus.

Electron micrographs of purified particles of miscanthus streak virus. Bars represent 100 nm (a) and 50 nm (b).

Parallel arrays of virus particles in the nucleus. Bar represents 250 nm.

Loosely aggregated virus particles (V) in the nucleus (N) and cytoplasm (C). Bar represents 500 nm.

Characteristic geometrical arrays of geminate particles in the vacuole. Bars represent 500 nm and 100 nm (inset).

Potential protein coding regions in the viral DNA (plus) strand (R) and its complementary (minus) strand (L). The coding capacity of each frame is given in kilodaltons. (Courtesy M. Ikegami.)
References list for DPV: Miscanthus streak virus (348)
- Bock, CMI/AAB Descr. Pl. Viruses 133, 4 pp., 1974.
- Bock & Harrison, AAB Descr. Pl. Viruses 297, 6 pp., 1985.
- Chatani, Mizuta, Shimizu, Furukawa & Ikegami, Abstr. 5th int. Congr. Pl. Path., Kyoto, Japan, Abstr. No. 2-137, p. 79, 1988.
- Francki & Hatta, CMI/AAB Descr. Pl. Viruses 221, 4 pp., 1980.
- Ikeda, Namba, Yamashita & Doi, Ann. phytopath. Soc. Japan 54: 79, 1988.
- Nonaka, Namba, Yamashita & Doi, Ann. phytopath. Soc. Japan 51: 58, 1985.
- Osaki & Inouye, CMI/AAB Descr. Pl. Viruses 232, 4 pp., 1981.
- Yamashita, Doi & Yora, Ann. phytopath. Soc. Japan 45: 128, 1979.
- Yamashita, Nonaka, Namba, Doi & Yora, Ann. phytopath. Soc. Japan 51: 582, 1985.