Details of DPV and References
DPV NO: 349 December 1989
Family: Pospiviroidae
Genus: Apscaviroid
Species: Apple scar skin viroid | Acronym: ASSVd
Apple scar skin viroid
H. Koganezawa Morioka Branch, Fruit Tree Research Station, Morioka 020-01, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described by Ohtsuka (1938) as ‘Manshu ringo sabi-ka byo’ (fruit russet disease of Manchurian apple), Millikan & Martin (1956) and Smith et al. (1956). The viroid was first described by Koganezawa et al. (1983).
Selected synonyms Dapple apple virus (Rev. appl. Mycol. 36: 327)
- Apple ‘sabi-ka’ virus (Rev. appl. Mycol. 44: 3089)
-
A viroid consisting of circular single-stranded RNA of 330 nucleotides. Transmitted mostly by graft-propagation and possibly by natural root grafting. Detected only in Malus and Pyrus. Widely distributed around the world. Common in China, where it is economically important, and in Japan.
Main Diseases
Causes severe scar skin (Fig. 1, Fig. 2, Fig. 5, Fig. 6), dappling (Fig. 4) or cracking on the surface of apple (Malus sylvestris) fruit. The entire crop from affected trees of susceptible cultivars becomes unsaleable.
Geographical Distribution
Widespread in Japan (Ushirozawa et al., 1968) and China (Liu et al., 1957). Also reported from Canada, France, Italy and USA (Kristensen, 1985).
Host Range and Symptomatology
Known hosts are confined to Malus and Pyrus in the Rosaceae (Liu et al., 1957). The viroid causes no symptoms in pear (Pyrus bretschneideri, P. serotina, P. ussuriensis and P. communis) (Liu et al., 1985). Transmitted experimentally by grafting and budding or by a razor-slash method. No infection was obtained by sap inoculation of 27 species of herbaceous plants in six families (P. Tien & W. Chen, personal communication).
-
Diagnostic species A wide range of Malus species and varieties respond with scar skin, dappling and cracking on the fruit, but differ in the type and severity of symptoms (Liu et al., 1957). The following apple (Malus sylvestris) cultivars are used in Japan and China.
- Cv. Indo. Fruits show scar skin only
(Fig. 1).
- Some sports of cv. Delicious (e.g. Starking Delicious). Dappling (Fig. 4) on the fruit 1 year after inoculation and scar skin (Fig. 2) some years later (Yamaguchi & Yanase, 1976).
- Cv. Ralls Janet. Scar skin with cracking on the fruit, and leaf rolling in young trees (Fig. 3). This leaf symptom, though recognized as a diagnostic symptom in China, is inconspicuous in Japan. This discrepancy may be ascribed to either a difference in the strains of viroid or a difference in the rootstocks used.
- The symptoms are usually concentrated near the calyx end of the fruit (Fig. 5, Fig. 6). When scar skin symptoms are mild, five streaks of scar tissue radiate from the calyx end (Fig. 5). For symptoms in other species and horticultural varieties of Malus, see Liu et al. (1957), Millikan (1963), McCrum et al. (1960), Stouffer (1983) and Ushirozawa et al. (1968).
- Some sports of cv. Delicious (e.g. Starking Delicious). Dappling (Fig. 4) on the fruit 1 year after inoculation and scar skin (Fig. 2) some years later (Yamaguchi & Yanase, 1976).
-
Propagation species Cultures may be propagated satisfactorily in most cultivars of Malus sylvestris (apple).
Assay species No suitable assay host is available. The viroid can be detected in bark tissues of apple trees by gel electrophoretic assay 1-2 years after inoculation.
Strains
Some isolates differ in the time of appearance and the severity of fruit scarring (Yamaguchi & Yanase, 1976). In one isolate from apple cv. American Summer Pearmain, the RNA has higher mobility than that of other strains in gel electrophoresis under non-denaturing conditions (H. Koganezawa, unpublished data).
Transmission by Vectors
No evidence for transmission by a vector. Field spread is believed to result from root contacts.
Transmission through Seed
No seed transmission was detected in apple (Ushirozawa et al., 1968).
Serology
No application of serological techniques has been reported.
Relationships
Apple scar skin viroid has 37% sequence homology with grapevine yellow speckle viroid. Both viroids share a common sequence in the central region of the molecule (Fig. 7), but lack the central conserved sequence of viroids in the potato spindle tuber viroid group. Other homologous residues also occur as blocks of base-paired residues in the secondary structures of apple scar skin and grapevine yellow speckle viroids. Apple scar skin viroid also has some homology with members of the potato spindle tuber viroid group but not with avocado sunblotch viroid (Koltunow & Rezaian, 1988).
Stability in Sap
No information.
Purification
The viroid is readily purified from bark or from young apple fruits by the following method (revised from Koganezawa, 1983 and Koganezawa, 1986). On a weight basis, the bark contains a higher concentration of the viroid than does the fruit. Pulverize the tissue in liquid nitrogen, and homogenize the powder in 2 vol 0.1 M Tris-HCl buffer, pH 8.5, 1 M NaCl, 1% SDS and 0.5% DIECA with polyvinylpolypyrollidone (1 g/10 g tissue) and chloroform (1 ml/10 g tissue). Centrifuge at low speed and precipitate the nucleic acid from the supernatant fluid with cold ethanol. Extract with phenol-chloroform. Remove polysaccharides by treating with 2-methoxyethanol and then precipitate the nucleic acids with cetyltrimethylammonium bromide. Fractionate by CF-11 cellulose column chromatography. Separate by two cycles of polyacrylamide gel electrophoresis, first under non-denaturing, then under denaturing conditions.
Properties of Infective Nucleic Acid
The molecule has M. Wt of 1.1 x 105 and contains 330 nucleotides (Fig. 7) with a base composition (G:A:C:U) of 29:20:31:20. The proposed secondary structure is a covalently closed single-stranded circular molecule, 68% base-paired, 61% of the base pairs are G:C, 29% A:U and 10% G:U (Hashimoto & Koganezawa, 1987).
Relations with Cells and Tissues
The viroid can be easily detected by polyacrylamide gel electrophoresis in extracts of bark and peel of infected apple trees. The concentration in leaves is low (Koganezawa, 1985; Saito et al., 1988).
Ecology and Control
In China, most pear trees are latently infected with the viroid. Mixed planting of apple and pear results in a high incidence of apple scar skin disease (Liu et al., 1957; Liu et al., 1985; Chen et al., 1988b).
Use of viroid-free propagating material is a main control measure. The agent, however, cannot be eliminated from apple trees by heat-treatment (Campbell & Sparks, 1976). Among the commercial apples, cvs Golden Delicious, Mutsu and Lord Lambourne are tolerant, and should not be used for topworking without prior indexing.
Notes
Dapple symptoms induced by the viroid in apple resemble those of apple fruit crinkle, a newly described disease (Koganezawa et al., 1989) with which a viroid-like RNA is also associated (Kanematsu et al., 1989). Apple fruit crinkle differs from apple scar skin in causing fruit crinkling in apple cv. Ohrin and in causing bark blister without fruit symptoms in apple cv. Starking Delicious. The mobility in gel electrophoresis of the associated circular RNA also distinguishes between these two diseases.
As well as apple scar skin viroid another RNA (ASSARNA-2) is associated with apple scar skin disease (Koganezawa, 1983). This RNA shows the properties of linear double-stranded RNA (Koganezawa, 1986), has no infectivity (H. Koganezawa, unpublished data) and does not hybridize with cDNA of apple scar skin viroid (Hashimoto & Koganezawa, 1987). In China, a further circular RNA (ASSDRNA-2) was found in diseased apple branches. ASSDRNA-2 has a higher molecular weight and is more stable thermodynamically than apple scar skin viroid (Chen et al., 1986; Chen et al., 1988a). The roles and functions of these secondary RNA species are unknown.
Figures

Apple cv. Indo: (left) healthy; (right) showing scar skin symptoms.

Apple cv. Starking Delicious: (left) healthy; (right) showing scar skin symptoms.

Apple cv. Ralls Janet showing leaf roll symptoms (courtesy of F. Liu).

Dapple symptoms on fruit of apple cv. Starking Delicious (courtesy of A. Yamaguchi).

Scar skin symptoms around the calyx end of apple fruit of cv. Starking Delicious.
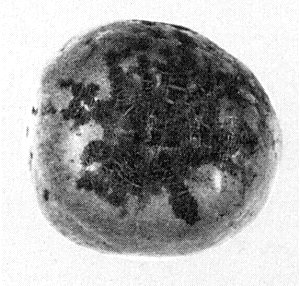
Scar skin symptoms around the calyx end of apple fruit of cv. Starking Delicious.
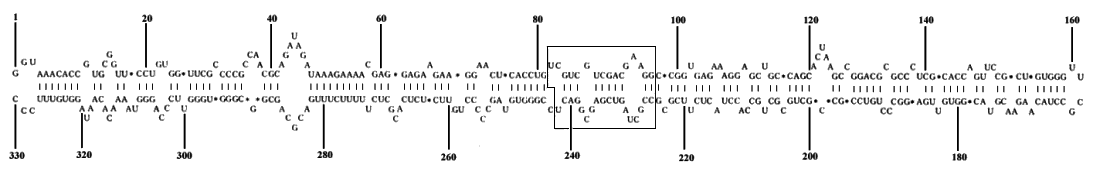
Nucleotide sequence of apple scar skin viroid (redrawn from Hashimoto & Koganezawa, 1987). The boxed area is the sequence common to apple scar skin viroid and grapevine yellow speckle viroid (Koltunow & Rezaian, 1988). In the nucleotide sequence (M36646) shown in the original paper (Hashimoto & Koganezawa, 1987) CU was mis-spelled as UC at nucleotides 228-229.
References list for DPV: Apple scar skin viroid (349)
- Campbell & Sparks Acta Hort. 67: 261, 1976.
- Chen, Lin, Tian, Liu, Wang & Wang, Chin. J. Virol. 4: 367, 1988a.
- Chen, Lin, Yang, Tien, Liu, Wang & Wang, Acta phytopath. sin. 18: 157, 1988b.
- Chen, Tian, Jin, Wang & Liu, Chin. J. Virol. 2: 366, 1986.
- Hashimoto & Koganezawa, Nucleic Acids Res. 15: 7045, 1987.
- Kanematsu, Koganezawa, Namba, Yamashita & Tsuchizaki, Ann. photopath. Soc. Japan 55: 545, 1989.
- Koganezawa, Bull. Fruit Tree Res. Stn, Japan C-10: 49, 1983.
- Koganezawa, Ann. phytopath. Soc. Japan 51: 176, 1985.
- Koganezawa, Acta Hort. 193: 29, 1986.
- Koganezawa, Yanase & Sakuma, Acta Hort. 130: 193, 1983.
- Koganezawa, Ohnuma, Sakuma & Yanase, Bull. Fruit Tree Res. Stn, Japan C-16: 57, 1989.
- Koltunow & Rezaian, Nucleic Acids Res. 16: 849, 1988.
- Kristensen, List of fruit tree virus and mycoplasma diseases, 1985, mimeographed listing prepared for Int. Soc. hort. Sci. Pl. Prot. Commission by Int. Committee for Cooperation in Fruit Tree Virus Research, 7 pp., 1985.
- Liu, Chen & Chen, Apple scar skin disease, Science Press, Beijing, 43 pp., 1957.
- Liu, Wang, Chen & Chen, China Fruit, 1985: 36, 1985.
- Millikan, in Tech. Commun. Commonw. Bur. Hort. Plantn Crops 30: 51, 1963.
- Millikan & Martin, Pl. Dis. Reptr 40: 229, 1956.
- McCrum, Barrat, Hilborn & Rich, Bull. Maine agric. Exp. Stn No. 595; Tech. Bull. New Hamps. agric. Exp. Stn No. 101: 63 pp., 1960.
- Ohtsuka, J. Jap. Soc. hort. Sci. 9: 282, 1938.
- Saito, Terauchi, Katsube, Yoshikawa & Takahashi, Abstr. 5th int. Congr. Pl. Path., 1988, Kyoto, Japan, p.71, 1988.
- Smith, Barrat & Rich, Pl. Dis. Reptr 40: 765, 1956.
- Stouffer, Acta Hort. 130: 141, 1983.
- Ushirozawa, Tojo, Takemae & Sekiguchi, Butt. Nagano hort. Res. Stn, Japan 7: 1, 1968.
- Yamaguchi & Yanase, Acta Hort. 67: 249, 1976.