Details of DPV and References
DPV NO: 352 December 1989
Family:
Genus:
Species: | Acronym:
Tombusvirus group
G. P. Martelli Dipartimento di Patologia vegetale, Università degli Studi and Centro di Studio del CNR sui Virus e le Virosi delle Colture Mediterranee, 70126 Bari, Italy
M. Russo Dipartimento di Patologia vegetale, Università degli Studi and Centro di Studio del CNR sui Virus e le Virosi delle Colture Mediterranee, 70126 Bari, Italy
D. Gallitelli Dipartimento di Patologia vegetale, Università degli Studi and Centro di Studio del CNR sui Virus e le Virosi delle Colture Mediterranee, 70126 Bari, Italy
Contents
- Type Member
- Main Characteristics
- Members
- Geographical Distribution
- Transmission by Vectors
- Ecology and Control
- Relations with Cells and Tissues
- Properties of Particles
- Genome Properties
- Replication
- Satellite
- Defective-Interfering RNA
- Relationships
- Notes on Tentative Members
- Affinities with Other Groups
- Notes
- Acknowledgements
- Figures
- References
Type Member
Tomato bushy stunt virus.
Main Characteristics
Isometric particles of a single type, c. 30 nm in diameter, which sediment at 127-136 S. Each particle is constructed of 180 subunits of one major protein species of M. Wt c. 40 x 103. The genome is a single molecule of positive sense ssRNA of M. Wt 1.5 x 106 (c. 4700 nucleotides), constituting c. 17% of the particle weight. Thermal inactivation point 80-90°C, longevity in sap 4-5 weeks, dilution end-point usually about or above 10-6, concentration in sap up to 150 mg virus per kg leaf tissue. Most tombusviruses have narrow natural but wide experimental host ranges; they tend to remain localized within artificially infected hosts. In infected cells, virus particles occur in the cytoplasm and vacuoles, sometimes within mitochondria and/or the nucleus. Infected cells contain cytopathic structures called multivesicular bodies, derived from altered peroxisomes or mitochondria. All members are readily transmitted by inoculation of sap and can also be acquired from the soil. One member has a fungal vector. Some are transmitted through seed.
Recent reviews on tombusviruses are by Martelli (1981), Francki et al. (1985) and Martelli et al. (1988).
Members
Definitive and tentative members of the group with some of their properties are:
ND = Not determined
Virus and acronym
Description No
or Ref.Sedi-
mentation
coefft
(s20,w)Buoyant
density
in CsCl
(g/cm3)M. Wt of
coat
protein
(kd)M. Wt of
RNA
(kb)Serological
relatedness
to
TBSV-typeOrigin of
multivesicular
bodies in
Chenopodium quinoa
A. DEFINITIVE MEMBERS
Artichoke mottled crinkle (AMCV)
69
132
1.35
43
4.70
+ + +
peroxisomes
Eggplant mottled crinkle (EMCV)
Makkouk et al., 1981
132
134
41
4.70
+
peroxisomes, vesicles in chloroplasts
Carnation Italian ringspot (CIRV)
69
135
136
37
4.70
+
mitochondria
Cucumber necrosis (CNV)
82
133
ND
40
4.70
-
ND
Cymbidium ringspot (CyRSV)
178
127
1.36
43
4.70
±
peroxisomes and mitochondria
Grapevine Algerian latent (GALV)
Gallitelli et al., 1989b
128
1.34
37
4.70
-
peroxisomes, vesicles in chloroplasts and mitochondria
Lato river (LRV)
Gallitelli et al., 1989a
ND
1.35
38
4.70
+
peroxisomes
Moroccan pepper (MPV)
Fischer & Lockhart, 1977;
Gallitelli & Russo, 1987
140
1.36
43
4.70
++
peroxisomes
Neckar river (NRV)
Koenig & Lesemann, 1985;
Gallitelli & Russo, 1987
133
1.36
45
4.70
+
peroxisomes, vesicles in chloroplasts and mitochondria
Petunia asteroid mosaic (PAMV)
69
134
1.35
41
4.70
+++
peroxisomes
Pelargonium leaf curl (PLCV)
69
136
1.34
41
4.70
+++
peroxisomes
Tomato bushy stunt (TBSV)
69
130
1.35
42
4.70
++++
peroxisomes, mitochondria
B. TENTATIVE MEMBERS
Cucumber leafspot (CLSV)
319
127
1.34
44
4.50
-
No multivesicular bodies formed
Cucumber soil- borne (CSBV)
Koenig et al., 1983
120
ND
41
4.50
-
No multivesicular bodies formed
Geographical Distribution
TBSV strains (strain BS-3 in particular) occur throughout Europe, the Mediterranean basin, and North and South America. PLCV and AMCV follow in part the distribution of their natural hosts, probably as a result of dissemination in vegetative planting material. PAMV is recorded in cherry and other woody hosts from several European countries and Canada (Koenig & Kunze, 1982). Records of MPV exist from Germany and Morocco (Vetten & Koenig, 1983) and of EMCV from Lebanon (Makkouk et al., 1981) and India (Raj et al., 1989). Other tombusviruses may have restricted distributions.
Transmission by Vectors
Tombusviruses are soil-borne, but except for CNV, their transmission does not seem to be mediated by living organisms. Repeated attempts to transmit TBSV and PAMV with the chytrid fungi Olpidium brassicae and Lagenocystis radicicola have failed (Teakle & Gold, 1963; Allen & Davidson, 1967; Campbell, 1969; Campbell et al., 1975). CNV is transmitted by Olpidium radicale. Virus particles, but not naked viral RNA, are adsorbed to the plasmalemma and the axonemal sheath of the zoospores, and taken up by endocytosis or, more commonly, flagellar retraction (Stobbs et al., 1982). No insect or mite vector is known (Lovisolo, 1957; Hollings, 1962; Orlob, 1968; Hollings et al., 1977).
Ecology and Control
Field spread of PAMV, PLCV and TBSV in their respective natural hosts has been observed in several countries (Lovisolo et al., 1965; Hollings & Stone, 1965; Cherif, 1981). These viruses, as well as other definitive members of the group (MPV, CyRSV) are established in soils, from which they can be acquired by host plants. Transplanting healthy seedlings into soils that either contained infected plants or were made infective by the addition of plant sap or concentrated virus suspensions, resulted in 10 to 100% infection, depending on the soil type and on the virus/host combination (Lovisolo et al., 1965; Hollings & Stone, 1965; Hollings et al., 1977; Kegler et al., 1980; Kegler & Kegler, 1981; Cherif, 1981; Vetten & Koenig, 1983; Tomlinson & Faithfull, 1984). In certain soils TBSV retains infectivity for up to 7 months and resists autoclaving at 121°C for 2 h (Kegler et al., 1980; Cherif, 1981). PAMV, TBSV and CyRSV have been recovered from leachates of soils containing infected plants (Campbell et al., 1975; Hollings et al., 1977; Cherif, 1981; Tomlinson & Faithfull, 1984; M. El Hamry & H. U. Fischer, personal communication). Furthermore, particles of the type and BS-3 strains of TBSV occur in the water of rivers and lakes of several countries (Tomlinson & Faithfull, 1984; Koenig & Lesemann, 1985). NRV was first discovered in the river Neckar in Germany (Koenig & Lesemann, 1985), LRV in the river Lato in southern Italy (Vovlas & Di Franco, 1987), and CIRV in a small stream in Germany (Buttner et al., 1987). According to Tomlinson & Faithfull (1984), TBSV may enter rivers from drainage water into which it is released from the roots of infected hosts or, more commonly, from sewage water which it reaches by passing unharmed through the alimentary tract of man (Tomlinson et al., 1982).
Although no seed transmission was found with PLCV, PAMV or CyRSV in naturally or artificially infected hosts (Hollings, 1962; Lovisolo et al., 1965; Hollings et al., 1977), infected seeds may constitute a route for dispersal of some tombusviruses. For TBSV the frequency of seed transmission was low (no more than 7%) in apple (Allen, 1969; Kegler & Schimanski, 1982), whereas in pepper and tomato it ranged from around 5% (Cherif, 1981) to more than 50% (Tomlinson & Faithfull, 1984). CyRSV is the only member of the group reported to be transmissible by foliage contact between plants (Hollings et al., 1977).
Relations with Cells and Tissues
Tombusviruses tend to remain localized in infected tissues, especially in artificially inoculated plants. With TBSV it was postulated that a flavonoid accumulating around and within the lesions induced by the virus may play a role in the localization of the infection (Redolfi et al., 1978). The cytopathology and histochemistry of local lesions induced in Gomphrena globosa by TBSV and CyRSV and the time course of events leading to ultrastructural modifications were found to be the same (Appiano et al., 1977, 1981; Pennazio et al., 1978; Russo & Martelli, 1981). Systemic invasion is more common in plants infected naturally, although certain members of the group (TBSV, PAMV, MPV) are not always readily translocated from roots, the site of infection, to the foliage (Lovisolo et al., 1965; Vetten & Koenig, 1983; Tomlinson & Faithfull, 1984).
Tombusviruses invade all tissues of systemically infected host plants. Symptoms, when expressed, are clear-cut and remain visible for the whole vegetative season. However, pelargonium and artichoke plants, infected with PLCV and AMCV respectively, undergo apparent ‘recovery’ in spring when temperatures rise (Hollings & Stone, 1965; Quacquarelli & Martelli, 1966).
The effects of tombusvirus infection on cells have been studied mainly in leaf parenchyma tissues with either localized or systemic infections. Inclusions visible with the light microscope after staining (e.g. Azure B) are induced in cells infected with TBSV (Christie & Edwardson, 1977). In thin-sectioned tissues, virus particles are found mainly in the cytoplasm, but also in the central vacuole and cell organelles (nuclei and mitochondria, rarely in chloroplasts). Virus particles may accumulate within bubble-like extensions of the tonoplast protruding into the central vacuole, or may aggregate in the cytoplasm to form amorphous or crystalline inclusions. Excess coat protein can also accumulate in the cytoplasm forming small electron-dense patches (Appiano et al., 1986; Russo et al., 1987). ‘Multivesicular bodies’ are another type of cytoplasmic inclusion characteristically induced by all definitive members of the group (Martelli et al., 1977, 1988; Russo et al., 1987), but not by the two tentative members CSBV and CLSV (Di Franco & Martelli, 1987). They are globose to ovoid structures made up of an outer single-membraned envelope lined with vesicles, which surround a central vacuole-like area. Depending on their origin, the multivesicular bodies may contain electron-dense material in granular or paracrystalline form, or remnants of mitochondrial stroma and cristae. The vesicles originate as invaginations of the boundary membrane. They contain dsRNA, possibly representing replicative forms or replicative intermediates of the viral genome (Russo et al., 1983; Di Franco et al., 1984). Depending on the virus and, to a lesser extent, on the host, multivesicular bodies may arise from deranged peroxisomes and/or mitochondria, but not from chloroplasts (Appiano & D'Agostino, 1983; Martelli et al., 1984; Di Franco et al., 1984; Bassi et al., 1986); they are also formed in protoplasts (De Varennes et al., 1984). Virus-specific cytopathic effects that allow group members to be differentiated from one another are observed in Chenopodium quinoa (Russo et al., 1987). These are: nuclear membraneous inclusions (AMCV); development of multivesicular bodies only from mitochondria (CIRV), or equally from peroxisomes and mitochondria (CyRSV), or from peroxisomes and occasionally from mitochondria (TBSV); development of multivesicular bodies from peroxisomes and vesiculation of some mitochondria and chloroplasts (GALV and NRV); development of multivesicular bodies from peroxisomes and extensive vesiculation of chloroplasts (EMCV). Furthermore, in G. globosa infected with the type and BS-3 strains of TBSV, the chloroplasts exhibit specific structural modifications and an apparently unique type of association with multivesicular bodies (Martelli et al., 1984).
Properties of Particles
Tombusvirus particles sediment as a single component at 127-140 S and are stable in CsCl in which their buoyant densities are 1.34-1.36 g/cm (Martelli et al., 1988). In electrophoresis the particles of all definitive members migrate as a single species. The rate and direction of migration, however, vary with individual members and are influenced by the conditions of electrophoresis, such as type of medium, type and molarity of buffer, and pH (Martelli et al., 1988).
The protein shell of tombusviruses is composed of 180 identical subunits with a M. Wt of c. 40,000 (range 37,000-45,000) arranged in a T = 3 icosahedral surface lattice. The subunits show a dimeric clustering at the surface of the particles giving rise to 90 morphological units located on the two-fold axes of the lattice (Finch et al., 1970; Crowther & Amos, 1971; Harrison et al., 1978). Minor polypeptides were detected, the smallest of which (M. Wt c. 28,000) may be a cleavage product of the major coat protein (Ziegler et al., 1974). A larger polypeptide (M. Wt c. 87,000) occurring in the proportion of one molecule per virus particle, formerly suggested to be a polymerase attached to the viral RNA (Butler, 1970), has now been identified as a stable covalently linked dimer of two protein coat subunits (Stockley et al., 1986). The coat protein subunits of TBSV contain 387 amino acid residues, whose sequence has been determined (Hopper et al., 1984).
Each coat protein subunit has four regions: an internal positively charged domain (R) comprising 66 residues which, together with an ‘arm’ 36 residues long, constitutes the polypeptide's N-terminal region. This region is flexibly linked to two distinct globular parts, of which one (domain S, 168 residues) forms the shell of the virus particle, and the other (domain P, 127 residues) protrudes on the surface of the particle (Harrison et al., 1978; Olson et al., 1983; Harrison, 1984; Hopper et al., 1984). The P domains are clustered in pairs, giving rise to the 90 projections that constitute the units contrasted by negative staining in the electron microscope. Because of their flexibility, the coat polypeptides can assume three distinct packing orientations designated A, B and C (for details see Harrison et al., 1978; Olson et al., 1983; Harrison, 1984). The connecting arms of the 60 subunits in orientation C are folded in an orderly manner at the bottom of their respective S domains, interlocking to form an internal framework which determines particle size (Olson et al., 1983). The N-termini of the remaining 120 subunits in orientations A and B and the positively charged residues of the inward-facing S domain surfaces are potential sites for RNA binding (Harrison et al., 1978; Chauvin et al., 1978; Hopper et al., 1984). Particle stability depends on the P domain dimer contacts, which are very stable, and on trimer interactions between the S domains of A, B and C subunits. The strength of these latter contacts is enhanced by Ca2+ ions (two for each pair of interacting subunits), but not by Mg2+ ions (Robinson & Harrison, 1982; Kruse et al., 1982; Hogle et al., 1983). Removal of Ca2+ at about pH 7 induces reversible swelling of the particles. These, unlike the swollen particles of bromoviruses (Bancroft, 1970) and sobemoviruses (Hsu et al., 1976) are not sensitive to RNase, but are partially sensitive to protease. The terminal regions of some subunits undergo proteolytic attack, possibly because they are extruded through gaps in the expanded shell (Robinson & Harrison, 1982; Kruse et al., 1982). From hydrogen-ion titration curves, it has been established that the protein shell of TBSV is semipermeable, thus having some of the ion-handling properties of lipid membranes (Durham et al., 1984). The genomic RNA is tightly packed within the protein shell, most of it being probably located in the space (c. 3 nm) between the S domains and the internal concentric shell made up of the N-terminal regions (Chauvin et al., 1978).
Upon prolonged storage at 5°C, purified preparations of LRV, but not of other members of the group (AMCV, CIRV, MPV, RNV, TBSV-type) give rise to particles c. 17 nm in diameter. These particles are not infective, do not contain nucleic acid, are made up of a single protein species with M. Wt of c. 27,000, and are serologically indistinguishable from ordinary LRV virions. They are thought to be formed by spontaneous reassembly of partially degraded coat proteins subunits (Gallitelli et al., 1989a).
Genome Properties
The genomic RNA (G-RNA) of tombusviruses has a chain lengh of c. 4.7 kilobases (kb) as estimated by electrophoresis in agarose gel under denaturing conditions (Gallitelli et al., 1985) and determined by sequence analysis: 4701 and 4733 bases, respectively, for CNV (Rochon & Tremaine, 1989) and CyRSV (Grieco et al., 1989a). Molar base ratios of AMCV, CyRSV, PAMV and AMCV RNA range between: G 27.8-28.6 : A 23.5-26.8 : C 21.1-21.5 : U 23.4-26.8 (Martelli, 1981). G-RNAs of TBSV and CyRSV do not have a covalently bound 5' terminal protein (VPg) necessary for infectivity (Mayo et al., 1982; Burgyan et al., 1986), but the 5' terminus is protected probably by a cap (Grieco et al., 1989a; Rochon & Tremaine, 1989). The 3' ends of the G-RNA molecules of CyRSV, CNV and PAMV do not possess polyadenylate sequences (Grieco et al., 1989a, 1989b; Rochon & Tremaine, 1989; Hillman et al., 1989). Dot blot hybridization analysis with complementary DNA (cDNA) to TBSV has shown that all tested tombusviruses have sequence homology with TBSV and among themselves (Gallitelli et al., 1985, 1989a, 1989b; Gallitelli & Russo, 1987; Koenig & Burgermeister, 1988; Rochon & Tremaine, 1989). Comparison of nucleotide sequence of CNV (Rochon & Tremaine, 1989), CyRSV (Grieco et al., 1989a, 1989b), PAMV (Hillman et al., 1989) and AMCV (F. Grieco & D. Gallitelli, unpublished information) has confirmed the existence of extensive sequence homology. In addition to G-RNA, two subgenomic RNA species of about 2.1 kb (sg1) and 0.9 kb (sg2) were found in plants infected with CyRSV, TBSV and PAMV (Russo et al., 1988; Hayes et al., 1988; Hillman et al., 1989). Both sg1 and sg2 are encapsidated in the particles of CyRSV (Russo et al., 1988) and TBSV (Hayes et al., 1988), whereas only sg1 is encapsidated in the particles of PAMV (Hillman et al., 1989). The occurrence of similar virus-related RNA species has been postulated for CNV (Rochon & Tremaine, 1989). G-RNA, sg1 and sg2 have the same 3' terminus; sg1 is translated in vitro into the virus coat protein (M. Wt c. 41 K), whereas sg2 is translated into a polypeptide of M. Wt c. 22 K (Russo et al., 1988; Grieco et al., 1989b; Hayes et al., 1988; Hillman et al., 1989; Rochon & Tremaine, 1989). A protein of M. Wt c. 19 K may also be translated from sg2 in a different reading frame, but has not been detected experimentally (Burgyan et al., 1986; Russo et al., 1988). Translation of G-RNA produces a polypeptide with an estimated M. Wt of 40 K for CyRSV (Burgyan et al., 1986) or 37 K for TBSV (Hayes et al., 1988). However, sequence analyses show that the first cistron in G-RNA encodes a protein of M. Wt c. 33 K terminating with an amber stop codon (Rochon & Tremaine, 1989; Grieco et al., 1989b). Readthrough of the amber codon would produce a polypeptide of M. Wt c 92 K. Such a readthrough product was detected in vitro using genomic TBSV RNA as a messenger, but only when calf liver tRNA was added to the rabbit reticulocyte lysate (Hayes et al., 1988). The only protein products detected in vitro were the coat and the 22 K proteins of CyRSV (Burgyan et al., 1986).
The genome organization of CyRSV is shown in Fig. 1; it applies also to CNV (Rochon & Tremaine, 1989) and, most probably, to all members of the group. The function of the 22 K protein is unknown since no homology was found between the CyRSV or CNV 22 K proteins and non-structural proteins of other unrelated viruses (Grieco et al., 1989b; Rochon & Tremaine, 1989). By contrast, the 92 K protein may enter the constitution of viral replicase as suggested by amino acid sequence similarities with putative replicases of other viruses (Grieco et al., 1989a; Rochon & Tremaine, 1989).
Replication
Little is known on the site of replication of tombusviruses. They may replicate in the cytoplasm, the mutivesicular bodies being possible sites of viral RNA synthesis. Both single-stranded and double-stranded replicative forms of CyRSV genomic and subgenomic RNAs have been isolated from infected plant tissues. In cowpea protoplasts inoculated with CyRSV, cordycepin and actinomycin D inhibited virus multiplication lowering both the proportion of protoplasts that became infected, and the virus yield per infected protoplast. No inhibition was obtained with chloramphenicol, which suggests that 70 S ribosomes are not involved in the synthesis of CyRSV coat protein (Russo & Gallitelli, 1985). G-RNA and two subgenomic RNAs were released by treatment with puromycin from polysomes extracted from TBSV-infected plants (Hayes et al., 1988), and double-stranded (ds) RNA species corresponding in length to the three major single-stranded (ss) RNA species were detected in plants infected with TBSV (Hayes et al., 1984), PAMV (Hillman et al., 1985), CNV (Rochon & Tremaine, 1987), and CyRSV (Russo et al., 1988). Although replication of G-RNA via the formation of full-length minus-strand RNA might be expected, it is unknown whether the subgenomic RNA species replicate in the same way, or are derived by internal initiation on the genomic minus-strand RNA.
Satellite
Most tombusviruses, in addition to G-RNA, have a linear molecule of satellite RNA (S-RNA) with an estimated chain length of 0.7 kb (Gallitelli & Hull, 1985). S-RNA is packaged in the protein shell of the helper virus, possibly in particles that do not contain G-RNA. As determined by Gallitelli & Hull (1985), the S-RNA species are not infective on their own, have extensive sequence homology (up to 83%) with one another but none with the G-RNA of the helper virus. However, recent studies have shown that the 5' region of CyRSV S-RNA has a sequence of 53 nucleotides exhibiting 88% homology with a comparable sequence in the 5' region of both the G-RNA and a defective-interfering RNA (L. Rubino, personal communication). The replication of S-RNA from TBSV-type strain is supported by the G-RNA molecules of other definitive tombusviruses. S-RNA from CyRSV has no messenger activity in cowpea protoplasts or in rabbit reticulocyte lysates (Burgyan et al., 1986). Studies on the replication of CyRSV S-RNA have shown that: (i), as with G-RNA, no polyadenylate sequences are present at the 3' end of the molecule, which has a free -OH; (ii), the 5' terminus does not have phosphate or free hydroxyl groups, but is protected, probably by a cap structure; (iii), the S-RNA molecule is linear; (iv), its replication is impaired above 35°C; (v), in infected plants, besides the standard molecule of 0.7 kb, there are at least three multimeric species occurring as single-stranded or double-stranded molecules (Burgyan & Russo, 1988). With most definitive tombusviruses, the appearance of S-RNA seems to be host-related because S-RNA is acquired by satellite-free isolates of the helper virus when passaged through Nicotiana benthamiana. No S-RNA was found in over 300 artichoke plants naturally infected with AMCV. However, wild AMCV isolates acquired S-RNA when inoculated to N. benthamiana (D. Gallitelli, unpublished information). The presence of S-RNA induces attenuation of the symptoms of the helper virus (Gallitelli & Hull, 1985).
Defective-Interfering RNA
A symptom-modulating RNA differing from S-RNA has been found associated with PAMV, and identified as a type of defective interfering RNA (DI-RNA) (Hillman et al., 1985, 1987). This DI-RNA is a linear molecule of c. 0.4 kb which represents a deletion mutant of the helper virus G-RNA (being made up of a mosaic of fragments derived from both extremities and some internal tracts of the viral genome) and is packaged in the protein shell of the helper virus. Its presence in the inoculum modulates the symptoms and interferes with virus synthesis, thus decreasing virus yields (Hillman et al., 1987). Another DI RNA has been found associated also with CyRSV infections. As with PAMV, it is a mosaic-type RNA modulating symptom expression of the helper virus more effectively than S-RNA (Burgyan et al., 1989).
Relationships
The serology of tombusviruses has been studied in detail by Hollings & Stone (1975) and Koenig & Gibbs (1986). Except for CNV, all other definitive members of the group are serologically interrelated. The relationships (Fig. 2) range from close (e.g. PAMV and AMCV, SDI = 1), to intermediate (e.g. TBSV-type and AMCV or PLCV, SDI = 3), to distant (e.g. TBSV-type and EMCV or CIRV; MPV and GALV, SDI = 5-7), to nearly undetectable (e.g. TBSV-type and CyRSV, SDI >9). This situation is complicated by the existence of serological variants regarded as strains of individual members, as with TBSV (see Martelli, 1981), PLCV (Hollings & Stone, 1975), CyRSV (Hollings et al., 1977) and AMCV (Rana & Kyriakopoulou, 1982). Thus, where very close relationships exist, such as in the cluster comprising TBSV, AMCV, PAMV and PLCV, it can be hard to decide whether isolates are best considered to be different viruses or different strains. The interrelationships among all definitive tombusviruses were confirmed by cDNA hybridization analysis, but a clear-cut ranking of individual members based on the extent of sequence homology of their G-RNA molecules has not always been possible (Gallitelli et al., 1985; Koenig & Burgermeister, 1988). The two tentative members of the group are not serologically related to one another, nor, apparently, to any of the definitive members (Koenig et al., 1983; Gallitelli et al., 1983; Weber & Stanarius, 1984).
Affinities with Other Groups
Besides tombusviruses, there are two additional taxonomic groups of plant viruses with isometric particles and monopartite ssRNA genomes of M. Wt 1.3-1.5 x 106, i.e. the sobemovirus group (Matthews, 1982) and the carmovirus group (type member, carnation mottle virus) (Morris & Carrington, 1988). Members of the tombusvirus and carmovirus groups have striking structural homologies in the capsid protein, with respect to their polypeptide folding topology and subunit interactions (Harrison et al., 1978; Hogle et al., 1986; Carrington et al., 1987), and have similar physico-chemical properties (Martelli et al., 1988; Morris & Carrington, 1988). Moreover, some carmoviruses, namely turnip crinkle, galinsoga mosaic, glycine mottle, hibiscus chlorotic ringspot and melon necrotic spot viruses, induce cytological changes (multivesicular bodies) resembling those of definitive tombusviruses (see Francki et al., 1985; Di Franco & Martelli, 1987; Russo et al., 1987). However, wide differences exist between tombusviruses and carmoviruses in biological properties (host range and ecology) with one noteworthy exception, i.e. both CNV and melon necrotic ringspot carmovirus have cucumber as the natural host and are transmitted by O. radicale (Stobbs et al., 1982; Hibi & Furuki, 1985).
The organization of the viral genome is different in the two groups, and these differences are the basis for separating the two groups. As compared with tombusviruses, carmoviruses have a smaller genomic RNA (3.5-4.0 kb instead of 4.7 kb), from which are generated two 3'-proximal subgenomic RNAs differing in size from those of tombusviruses (1.5 and 1.7 kb instead of 0.9 and 2.1 kb) (Morris & Carrington, 1988); the electrophoretic patterns of dsRNA preparations of members of the two groups are also distinct (Rochon & Tremaine, 1987). The coat protein gene of carmoviruses (CarMV and TCV), unlike that of the tombusviruses (PAMV, CyRSV and CNV), is located at the 3' end of the genome and is translated from the smaller of the two subgenomic RNAs (Morris & Carrington, 1988).
Affinities with sobemoviruses are confined to the architectural organization of the protein shell, which appears to be very similar in the two groups (Abad-Zapatero et al., 1980; Harrison, 1984).
Figures

Genome organization of CyRSV. A. Relative position of the open reading frames (ORF) on the genomic RNA molecule. B. Translation products of the different genes expressed as molecular weights of the encoded proteins. C. Diagrammatic representation of the replication strategy of the virus: 33K and 92K proteins (possible components of the viral replicase) are expressed through genome-sized (G) template translation. Coat protein (41K), 22K, and 19K proteins are expressed through formation of, and subsequent translation from, subgenomic RNAs (sg1 and sg2).
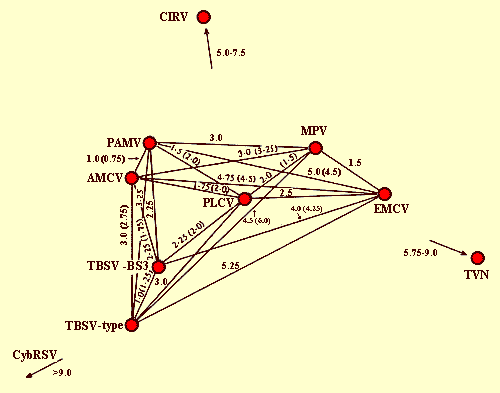
Serological classification of definitive tombusviruses (all included except for CNV, GALV and LRV) based on the average serological differentiation indices from reciprocal tests (RT-SDI), depicted as length units. With some viruses, there are slight discrepancies between calculated RT-SDI and the distances shown which, in these instances, are given in parentheses (from Koenig & Gibbs (1986)).
References list for DPV: Tombusvirus group (352)
- Abad-Zapatero, Abdel-Mesquid, Johnson, Leslie, Rayment, Rossmann, Suck & Tsukihara, Nature, Lond. 286: 33, 1980.
- Allen, Can. J. Pl. Sci. 49: 797, 1969.
- Allen & Davidson, Can. J. Bot. 4: 2375, 1967.
- Appiano & D'Agostino, J. Ultrastruct. Res. 85: 239, 1983.
- Appiano, Pennazio, D'Agostino & Redolfi, Physiol. Pl. Path. 11: 327, 1977.
- Appiano, D'Agostino, Redolfi & Pennazio, J. Ultrastruct. Res. 76: 173, 1981.
- Appiano, D'Agostino, Bassi, Barbieri, Viale & Dell'Orto, J. Ultrastruct. molec. Struct. Res. 97: 31, 1986.
- Bancroft, Adv. Virus Res. 16: 99, 1970.
- Bassi, Barbieri, Appiano & D'Agostino, J. Ultrastruct. molec. Struct. Res. 96: 194, 1986.
- Burgyan & Russo, J. gen. Virol. 69: 3089, 1988.
- Burgyan, Russo & Gallitelli, J. gen. Virol. 67: 1149, 1986.
- Burgyan, Grieco & Russo, J. gen. Viral. 70: 235, 1989.
- Butler, J. molec. Biol. 52: 589, 1970.
- Buttner, Jacobi & Koenig, J. Phytopath. 118: 131, 1987.
- Campbell, Pl. Dis. Reptr 52: 589, 1969.
- Campbell, Lovisolo & Lisa, Phytopath. Mediterranea 14: 82, 1975.
- Carrington, Morris, Stockley & Harrison, J. molec. Biol. 194: 265, 1987.
- Chauvin, Witz & Jacrot, J. molec. biol. 124: 641, 1978.
- Cherif, Thèse Diplome Docteur 3eme Cycle, Univ. P. & M. Curie, Paris, 84 pp., 1981.
- Christie & Edwardson, Monogr. Ser. Fla agric. Exp. Stn No. 9, 150 pp., 1977.
- Crowther & Amos, Cold Spring Harbor Symp. Quant. Biol. No. 36: 489, 1971.
- De Varennes, Russo & Maule, J. gen. Virol. 65: 1851, 1984.
- Di Franco & Martelli, J. submicroscop. Cytol. 19: 605, 1987.
- Di Franco, Russo & Martelli, J. gen. Virol. 65: 1233, 1984.
- Durham, Witz & Bancroft, Virology 133: 1, 1984.
- Finch, Klug & Leberman, J. molec. Biol. 50: 215, 1970.
- Fischer & Lockhart, Phytopathology 67: 1352, 1977.
- Francki, Milne & Hatta, An Atlas of Plant Viruses Vol. 1. Boca Raton: CRC Press, 222 pp., 1985.
- Gallitelli & Hull, J. gen. Virol. 66: 1533, 1985.
- Gallitelli & Russo, J. Phytopathol. 119: 106, 1987.
- Gallitelli, Vovlas & Avgelis, Phytopath. Z. 106: 149, 1983.
- Gallitelli, Hull & Koenig, J. gen. Virol. 66: 1523, 1985.
- Gallitelli, Di Franco, Vovlas & Martelli, J. Phytopath. 126: 213, 1989a.
- Gallitelli, Martelli & Di Franco, Phytoparasitica 17: 61, 1989b.
- Grieco, Burgyan & Russo, Nucleic Acids Res. 17: 6383, 1989a.
- Grieco, Burgyan & Russo, J. gen. Virol. 70: 2533, 1989b.
- Harrison, Trends biochem. Sci. 9: 345, 1984.
- Harrison, Olson, Schutt, Winkler & Bricogne, Nature, Lond. 276: 368, 1978.
- Hayes, Buck & Brunt, J. gen. Virol. 65: 1239, 1984.
- Hayes, Brunt & Buck, J. gen. Virol. 69: 3047, 1988.
- Hibi & Furuki, CMI/AAB Descr. Pl. Viruses 302, 4 pp., 1985.
- Hillman, Morris & Schlegel, Phytopathology 75: 361, 1985.
- Hillman, Carrington & Morris, Cell 51: 427, 1987.
- Hillman, Hearne, Rochon & Morris, Virology 169: 42, 1989.
- Hogle, Kirchhausen & Harrison, J. molec. Biol. 171: 95, 1983.
- Hogle, Maeda & Harrison, J. molec. Biol. 191: 625, 1986.
- Hollings, Ann. appl. Biol. 50: 189, 1962.
- Hollings & Stone, Ann. appl. Biol 56: 87, 1965.
- Hollings & Stone, Ann. appl. Biol 80: 37, 1975.
- Hollings, Stone & Barton, Ann. appl. Biol. 85: 233, 1977.
- Hopper, Harrison & Sauer, J. molec. Biol. 177: 701, 1984.
- Hsu, Seghal & Pickett, Virology 69: 587, 1976.
- Kegler & Kegler, Arch. Phytopath. PflSchutz 17: 307, 1981.
- Kegler & Schimanski, Arch. Phytopath. PflSchutz 18: 105, 1982.
- Kegler, Kleinhempel & Kegler, Arch. Phytopath. PflSchutz 16: 73, 1980.
- Koenig & Gibbs, J. gen. Virol 67: 75, 1986.
- Koenig & Burgermeister, J. Virol. Meth. 19: 57, 1988.
- Koenig & Kunze, Phytopath. Z. 103: 361, 1982.
- Koenig & Lesemann, Phytopath. Z. 112: 106, 1985.
- Koenig, Lesemann, Huth & Makkouk, Phytopathology 73: 515, 1983.
- Kruse, Kruse, Witz, Chauvin & Jacrot, J. molec. Biol. 162: 393, 1982.
- Lovisolo, Boll. Staz. Patol. veg. Roma, Ser. 111 14: 103, 1957.
- Lovisolo, Bode & Volk, Phytopath. Z. 53: 323, 1965.
- Makkouk, Koenig & Lesemann, Phytopathology 71: 572, 1981.
- Martelli, in Handbook of Plant Virus Infections and Comparative Diagnosis, p. 61, ed. E. Kurstak, Amsterdam: Elsevier/North-Holland, 943 pp., 1981.
- Martelli, Russo & Quacquarelli, in The Atlas of Insect and Plant Viruses, p. 257, ed. K. Maramorosch, New York: Academic Press, 478 pp., 1977.
- Martelli, Di Franco & Russo, J. Ultrastruct. Res 88: 275, 1984.
- Martelli, Gallitelli & Russo, in The Plant Viruses Vol. 3. Polyhedral Virions with Monopartite RNA Genomes, p. 13, ed. R. Koenig. New York: Plenum Press, 294 pp., 1988.
- Matthews, Intervirology 17: 1, 1982.
- Mayo, Barker & Harrison, J. gen. Virol. 59: 149, 1982.
- Morris & Carrington, in The Plant Viruses Vol. 3. Polyhedral Virions with Monopartite RNA Genomes, p. 73, ed. R. Koenig, New York: Plenum Press, 294 pp., 1988.
- Olson, Harrison & Bricogne, J. molec. Biol. 171: 61, 1983.
- Orlob, Virology 35: 121, 1968.
- Pennazio, Redolfi & Appiano, Inftore bot. Ital. 10: 363, 1978.
- Quacquarelli & Martelli, Atti 1° Congr. Un. Fitopatol. Mediterranea, Bari, 1966 Vol. 1: 68, 1966.
- Rana & Kyriakopoulou, Phytopath. Mediterranea 21: 101, 1982.
- Raj, Aslam, Srivastava & Singh, J. Phytopath. 125: 283, 1989.
- Redolfi, Cantisani & Pennazio, Phytopath. Z. 93: 325, 1978.
- Robinson & Harrison, Nature, Lond. 297: 563, 1982.
- Rochon & Tremaine, Abstr. 7th int. Congr. Virol., Edmonton, 1987: 85, 1987.
- Rochon & Tremaine. Virology 169: 251, 1989.
- Russo & Gallitelli, J. gen. Virol. 66: 2033, 1985.
- Russo & Martelli, J. Ultrastruct. Res. 77: 105, 1981.
- Russo, Di Franco & Martelli, J. Ultrastruct. Res. 82: 52, 1983.
- Russo, Di Franco & Martelli, Intervirology 28: 134, 1987.
- Russo, Burgyan, Carrington, Hillman & Morris, J. gen. Virol. 69: 401, 1988.
- Stobbs, Cross & Manocha, Can. J. Pl. Path. 4: 134, 1982.
- Stockley, Kirsh, Chow, Smart & Harrison, J. molec. Biol. 191: 721, 1986.
- Teakle & Gold, Virology 19: 310, 1963.
- Tomlinson & Faithfull, Ann. appl. Biol. 104: 485, 1984.
- Tomlinson, Faithfull, Flewett & Beard, Nature, Lond. 300: 637, 1982.
- Vetten & Koenig, Phytopath. Z. 108: 215, 1983.
- Vovlas & Di Franco, Inftore fitopatol. 37: 55, 1987.
- Weber & Stanarius, Arch. Phytopath. PflSchutz 20: 447, 1984.
- Ziegler, Harrison & Leberman, Virology 59: 509, 1974.