Details of DPV and References
DPV NO: 355 September 1998
Family: Unallocated ssRNA+ viruses
Genus: Umbravirus
Species: Groundnut rosette virus | Acronym: GRV
Groundnut rosette virus
A. F. Murant Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, Scotland, UK
D. J. Robinson Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, Scotland, UK
M. E. Taliansky Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, Scotland, UK
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Disease described first by
Zimmermann (1907) and subsequently by
Storey & Bottomley (1928) and
Storey & Ryland (1955,
1957).
Causal agent
first characterized by
Reddy et al. (1985a,
1985b). For
reviews of groundnut rosette disease and its causal agents see
Murant (1990b),
Murant et al. (1993), and
Robinson & Taliansky (1997).
Selected synonyms
Groundnut Kraüselkrankheit virus (Zimmermann, 1907; Rev appl. Mycol. 36: 303)
- Peanut rosette virus (Rev appl. Mycol. 19: 229)
Groundnut rosette virus (GRV) is a replicating ssRNA which does not produce a coat protein and therefore has no conventional particles. It depends on groundnut rosette assistor virus (GRAV; Luteoviridae) for encapsidation in GRAV coat protein and for transmission by Aphis craccivora in the persistent (circulative, non-propagative) manner. GRV supports the replication of a satellite RNA which is responsible for the symptoms of rosette disease in groundnut and plays an essential role in mediating the dependence of GRV on GRAV for transmission by aphids. GRV and its satellite RNA are manually transmissible to a small range of hosts. The virus complex occurs only in sub-Saharan Africa.
Main Diseases
Rosette is the most destructive virus disease of groundnut (peanut; Arachis
hypogaea) in Africa. In some years losses may be small, but in others
rosette epidemics can be responsible for devastating losses. Thus in 1975
rosette affected about 0.7 million ha of groundnut in Nigeria and caused yield
losses estimated at over 0.5 million tonnes, with a value estimated at US$ 250
million
(Yaycock et al., 1976).
In 1995, about 43,000 ha were affected in
eastern Zambia, with losses amounting to US$ 5 million
(Anon., 1996).
The
unpredictability of outbreaks is an important feature of the disease: the sudden
unexpected loss of an important source of protein and cooking oil, of income,
and of the seed for the next crop, may lead farmers to abandon growing
groundnuts in subsequent years. Following the epidemic in Malawi in 1994/5,
the area under groundnut decreased by 23%, from 89,000 to 69,700 ha
(Anon., 1996).
Rosette disease is caused by a complex of groundnut rosette virus (GRV),
together with an associated satellite RNA, and a helper virus,
groundnut rosette assistor virus
(GRAV; family Luteoviridae), on which the other two
components depend for transmission by the aphid Aphis craccivora.
Neither GRV nor GRAV themselves cause obvious symptoms in groundnut.
Murant & Kumar (1990)
showed that the disease symptoms are caused by
the GRV satellite RNA, different variants of the satellite being responsible for the
different major forms of rosette disease: chlorotic rosette
(Storey & Bottomley, 1928)
and green rosette
(Hayes, 1932;
Smartt, 1961;
Hull & Adams, 1968).
‘Mosaic rosette’
(Storey & Ryland, 1957)
is caused by
mixed infection with the ‘chlorotic’ satellite variant
and a ‘mottle’ variant
(Murant & Kumar, 1990).
The satellite RNA plays an essential role, as yet unexplained, in mediating the
dependence of GRV on GRAV for transmission by aphids
(Murant, 1990a).
This presumably explains why no satellite-free isolates of GRV
have been found in nature, though they can be obtained experimentally (see
below).
Plants with chlorotic rosette (Fig. 1, Fig. 2, Fig. 3) show a bright chlorosis of the leaves, usually with a few green islands. The chlorosis may affect the whole plant, or only some shoots or parts of shoots. Plants that are infected early are stunted, with small, curled and puckered leaflets. In the mosaic form of rosette (Fig. 4) the green parts of leaves are more extensive. In green rosette (Fig. 5), the leaves are very dark green, or show a light green and dark green mosaic, and are much reduced in size, with their margins rolled downwards. Plants that are infected early are severely stunted, with much shortened internodes, and appear as small dark green bushes. In all types of rosette, early infection causes severe or total loss of yield. Late infection can cause a decrease in the number and size of pods.
Geographical Distribution
Chlorotic rosette disease occurs throughout Africa south of the Sahara. Green rosette disease is found in West Africa and Uganda, and has recently been found in Angola (P. Subrahmanyam, personal communication), Malawi (Subrahmanyam & Mamba, 1993) and Swaziland (Subrahmanyam & Chiyembekeza, 1995). Mosaic rosette is reported only from East Africa. Groundnut rosette disease does not occur in other parts of the world; reports from Argentina, Australia, Fiji, India, Indonesia, Philippines and Russia have not been confirmed and are now discounted.
Host Range and Symptomatology
Experimentally, GRV, along with
GRAV
and any associated satellite RNA
species is transmissible from groundnut to groundnut by grafting. GRV and its
satellite RNA, but not GRAV, can be transmitted by manual inoculation to a
limited range of dicotyledonous plants, though manual inoculation to groundnut
(Arachis hypogaea) may be difficult, especially under greenhouse
conditions in summer. A procedure for efficient manual inoculation of GRV to
groundnut was reported by
Olorunju et al. (1992, 1995).
GRV has been
transmitted to several other species of Leguminosae (Glycine max, Indigofera
nummularifolia, Macrotyloma uniflorus, Phaseolus vulgaris, Stylosanthes
gracilis*, S. guayensis, S. mucronata*, S. juncea, S. sundaica*, Tephrosia
purpurea, Trifolium incarnatum*, Trifolium repens* and Vigna gracilis)
and to a few species in the Amaranthaceae (Gomphrena globosa*),
Chenopodiaceae (Chenopodium amaranticolor, C. murale, C.
quinoa, Spinacia oleracea*) and Solanaceae (Nicotiana
benthamiana, N. clevelandii, N. debneyi, N.occidentalis,
N. rustica, N. tabacum Samsun NN)
(Okusanya & Watson, 1966;
Adams, 1967;
Hull & Adams, 1968;
Dubern, 1980;
Reddy et al., 1985a;
Rajeshwari & Murant, 1988;
Kumar et al. 1991;
D. B. Dangora, personal communication). The plants marked * are also hosts of
GRAV.
Diagnostic species
- C. amaranticolor
. GRV cultures, with or without the satellite RNA, give minute necrotic lesions on inoculated leaves (Fig. 6) about 4 days after inoculation.
-
N. benthamiana. This is the most sensitive test plant. On inoculated
leaves, most GRV cultures, with or without the satellite RNA, give symptoms
ranging from almost none to necrotic spots, rings or target spots
(Fig. 7).
About
7-10 days after inoculation, veinal chlorosis or necrosis appears on the first
systemically infected leaf, accompanied usually by slight down-rolling. Later
this leaf often shows necrotic spots or line-patterns
(Fig. 7).
Leaves produced
subsequently usually show no more than a mild chlorotic mottle or they may be
symptomlessly infected. GRV cultures containing the yellow blotch satellite RNA
(Kumar et al., 1991)
induce symptoms similar to those of other isolates in
inoculated leaves but systemically infected leaves show a brilliant yellow blotch
mosaic affecting the entire plant and persisting for its lifetime
(Fig. 8). With all
isolates there is stunting of the plant.
- N. clevelandii (Fig. 9). Isolates with or without satellite RNA induce local chlorotic or necrotic spots or rings followed by systemic chlorosis or necrosis beginning about 7-10 days after inoculation. Later there is systemic mottle or mosaic, accompanied by crinkling and distortion of leaves. These symptoms eventually become less severe but the plant remains stunted. Cultures containing the yellow blotch satellite do not induce distinctive symptoms in this species.
Assay species
- C. amaranticolor
is the best local lesion host. Arachis hypogaea (groundnut) is used as a test plant in aphid transmission experiments.
- N. clevelandii (Fig. 9). Isolates with or without satellite RNA induce local chlorotic or necrotic spots or rings followed by systemic chlorosis or necrosis beginning about 7-10 days after inoculation. Later there is systemic mottle or mosaic, accompanied by crinkling and distortion of leaves. These symptoms eventually become less severe but the plant remains stunted. Cultures containing the yellow blotch satellite do not induce distinctive symptoms in this species.
Propagation species
- N. benthamiana
is the best propagation host for studies on virus properties and purification. For long term maintenance, virus cultures are best kept in Arachis hypogaea (groundnut).
Strains
No strains of GRV have been distinguished, but numerous variants of the satellite RNA have been described that affect the symptoms induced by GRV in plants; for example, different satellite variants are responsible for the chlorotic and green forms of rosette disease (Murant & Kumar, 1990). The mosaic form of rosette is caused by GRV containing a mixture of the ‘chlorotic’ satellite variant and a ‘mottle’ variant (Murant & Kumar, 1990). A ‘yellow blotch’ satellite variant discovered in the laboratory (Kumar et al., 1991) is responsible for a striking bright yellow symptom in N. benthamiana which has been useful as an experimental marker. Several satellite variants have been discovered experimentally that induce few or no symptoms in groundnut. One of these down-regulates the replication of GRV (Taliansky & Robinson, 1997a) and, for this reason, isolates containing it are difficult to maintain in culture.
Transmission by Vectors
The aphid Aphis craccivora is the only vector of any importance,
though there is a
single report of transmission by A. gossypii
(Adams, 1966).
However,
A. craccivora can transmit GRV and its satellite RNA only from source
plants that are also infected with
GRAV.
This is because GRV RNA does not
encode a coat protein, and relies on the GRAV coat protein for encapsidation
(A.F. Murant, unpublished data) and therefore for aphid transmission. A further
complication is that the satellite too must be present in the source plants for
GRAV-dependent transmission of GRV to occur, i.e. aphids transmit only GRAV
from plants containing GRAV plus a satellite-free culture of GRV
(Murant, 1990a).
The explanation for this is not known.
The dependence of GRV on GRAV is for acquisition and inoculation by the
aphid, not for infection of the inoculated plant. Therefore, in circumstances in
which the frequency of transmission of the two viruses is less than 100%, some
inoculated plants may become infected with only one of the viruses. Plants that
become infected with GRV alone cannot then serve as sources for aphid
transmission.
The GRAV/GRV/satellite complex is transmitted by A. craccivora in
the persistent (circulative, non-propagative) manner, the aphids retaining the
ability to transmit for at least 15 days, possibly for life
(Storey & Ryland, 1955;
Watson & Okusanya, 1967;
Dubern, 1980;
Misari et al., 1988).
Dubern (1980)
found a minimum acquisition access time of 4.5 h, a latent period
of 18 h, and a minimum inoculation access period of 3 min.
Misari et al. (1988),
working with separate cultures of chlorotic rosette and green rosette,
reported minimum acquisition access periods of 4 h and 8 h respectively, and
median latent periods of 26.4 h and 38.4 h respectively; the minimum inoculation
access period was 10 min for both cultures.
In all studies of vector relations reported so far, transmission has been assessed
by the appearance of rosette symptoms in the inoculated plants, so that the data
are strictly applicable only to GRV plus its satellite. However, the minimum
acquisition access times and latent periods for GRV are probably those of
GRAV. The minimum inoculation access time for GRV may well be shorter than
that for GRAV because GRV can infect mesophyll cells whereas GRAV needs to
be inoculated into the phloem.
Watson & Okusanya (1967) reported that A. craccivora populations from Nigeria and Kenya differed in ability to transmit cultures of the rosette virus complex from East and West Africa, but A. F. Murant (unpublished data) found that a population from Malawi transmitted GRAV isolates from Malawi and Nigeria.
Transmission through Seed
There is no evidence for transmission through seed or pollen.
Serology
GRV does not produce a coat protein, but a polyclonal antiserum has been raised against the non-structural 28K ‘movement’ protein (the product of ORF4) by injecting a rabbit with a fusion protein expressed in E. coli. The antiserum reacts with movement protein transiently expressed in N. benthamiana from a potato virus X (PVX)-based vector, but has not been shown to react with extracts from GRV-infected plants (E.V. Ryabov, unpublished data).
Relationships
GRV is a member of the genus Umbravirus
(Murant et al., 1995).
This genus comprises several imperfectly characterized ssRNA viruses which
lack coat protein genes and depend on unrelated helper viruses, usually
members of the family Luteoviridae, for encapsidation and for
transmission by aphids in a persistent (circulative, non-propagative) manner.
The genus Umbravirus is not assigned to a family but its closest affinities
seem to be with the family Tombusviridae.
Comparison of nucleotide sequences of genomic RNA molecules and of amino
acid sequences of proteins putatively encoded by the same RNA molecules
(Taliansky et al., 1996)
revealed close homologies between GRV and two
other viruses now classified in the genus Umbravirus,
carrot mottle mimic virus (syn. Australian isolate of
carrot mottle virus;
Gibbs et al., 1996a,
1996b)
and pea enation mosaic virus-2 (formerly the RNA-
2 of
pea enation mosaic virus;
Mayo & D'Arcy, 1998).
The sequences of GRV satellite RNAs have up to 59% homology with the satellite RNA associated with pea enation mosaic virus-2 (Demler et al., 1996).
Stability in Sap
(Reddy et al., 1985b). In 0.01 M Tris buffer, pH 8.0, containing 0.02 M sodium sulphite, much GRV infectivity survived after 1 day at room temperature or after 3 days at 4°C. A trace of infectivity survived even after 15 days at 4°C. Incorporation of 1 g/l Mg-bentonite did not affect these results. Buffer extracts were infective after dilution to 10-3 but not 10-4. Infectivity was abolished by treating the extracts with ether (50%), chloroform (10 or 50%), or butan-1-ol (8%).
Purification
Preparations of the infective ssRNA of GRV have not been obtained free from
host RNA, and no infection-specific band can be detected by electrophoresis of
whole RNA extracts from infected plants. However, unlike healthy plants,
infected plants yield abundant double-stranded RNA (dsRNA) which gives a
characteristic pattern of electrophoretic bands
(Reddy et al., 1985b;
Murant et al., 1988;
Murant & Kumar, 1990),
with
three characteristic dsRNA species
(Fig. 10).
Two of these species, dsRNA-1
(4.0 kbp) and dsRNA-2 (1.3 kbp), appear to be double-stranded forms of the
genomic and a sub-genomic RNA respectively. The third and much the most
abundant species, dsRNA-3 (0.9 kbp), is a double-stranded form of the satellite
RNA, from which GRV cultures can be freed experimentally
(Fig. 10)
(see
below). The characteristic dsRNA band pattern is a useful diagnostic aid and
Breyel et al. (1988)
described a quick procedure for analysing multiple
samples electrophoretically. However, somewhat similar dsRNA band patterns
are given by other putative umbraviruses (A.F. Murant, unpublished data), so
that such analyses should be interpreted with caution.
Preparation of infective ssRNA
The following procedure was described by
Murant et al. (1988):
1. Grind fresh leaf tissue (10 g) to a fine powder in liquid nitrogen with a pestle
and mortar. Stir the powder for 30 min at room temperature with 20 ml TSE
buffer (50 mM Tris-HCl, 0.1 M NaCl, 1 mM disodium EDTA, pH 7.0), 3.0 ml 10%
SDS, 40 mg bentonite and 30 ml ‘phenol reagent’ (9 vol. water-saturated phenol
+ 1 vol. m-cresol, containing 0.1% 8-hydroxy-quinoline).
2. Precipitate total nucleic acids from the aqueous phase with 70% ethanol and
place at -20°C for at least 16 h, then wash three times with cold 70%
ethanol. For infectivity assay, resuspend the pellets in 0.1 M phosphate buffer
pH 7.4 containing 1 mg/ml bentonite, and inoculate immediately to test plants.
Preparation of dsRNA
The following procedure was described by
Murant et al. (1988):
1. Make preparations of total nucleic acid as described above but after the
phenol extraction step adjust the aqueous phase to 20% (v/v) ethanol. Apply to
columns of Whatman CF-11 cellulose as described by
Dodds & Bar-Joseph (1983),
and elute the dsRNA fraction with ethanol-free TSE buffer. Recover
nucleic acid from the eluate by ethanol precipitation.
2. To remove any contaminating DNA, resuspend the sample in 50 mM Tris-HCl, 50
mM NaCl, 10 mM MgCl2, pH 7.5, and treat with 10 µg/ml
DNase I for 30 min at 30°C. Then, to remove contaminating
ssRNA, adjust to 0.3 M NaCl and treat with 10 ng/ml RNase A for 1 h at
30°C.
3. Adjust the preparation to 2% SDS and re-extract with ‘phenol reagent’.
4. A procedure for electrophoretic analysis of dsRNA preparations is given by
Murant et al. (1988).
Preparation of satellite-free cultures of GRV
The method used by
Murant & Kumar (1990) was as follows:
1. Electrophorese dsRNA preparations in 0.9% low-gelling-temperature agarose
gels and excise the dsRNA-1 bands from several tracks.
2. Boil the gel slices for 5 min in 0.5 ml 10 mM Tris-HCl, 1 mM EDTA, pH 7.4, then
cool rapidly on ice, add 4 mg/ml bentonite, and inoculate the preparation both
undiluted and at a dilution of 1/10 to Nicotiana benthamiana. Isolates
obtained from the highest infective dilution usually lack dsRNA-3 but possess
dsRNA-1 and dsRNA-2.
3. Biologically active preparations of dsRNA-3 are prepared and inoculated in
the same way, but of course the satellite does not replicate in the plants unless
they are also inoculated with melted dsRNA-1. Unlike dsRNA-1, dsRNA-3 does
not need to be melted to yield biological activity
(Kumar et al., 1991).
Particle Structure
No virus-like particles have been seen in preparations from plants infected only with GRV, and no candidate for the role of a particle protein has been identified among the proteins putatively encoded by the genomic RNA. The virus seems to exist in plants as an infective ssRNA. However, its relative stability in crude sap suggests that it is protected in some way against degradation by ribonucleases.
Genome Properties
The GRV genome is a single segment of
ssRNA. The complete nucleotide sequence has been determined
(Taliansky et al., 1996);
its mol. wt is 1.3 × 106 (4019
nucleotides). The sequences of ten variants of the satellite RNA were
determined by
Blok et al. (1994);
their nucleotide numbers ranged from
895 to 903. Radioactive and non-radioactive cDNA probes complementary to
the satellite RNA are useful for detection of GRV infection
(Blok et al., 1995).
Probes complementary to the GRV genomic RNA are also available but
seem likely to be less useful because of the lower molar concentration of
genomic RNA than of the satellite RNA in infected plants.
Neither the genomic RNA nor the satellite RNA are polyadenylated, nor do either
of them possess a 3'-terminal tRNA-like sequence. It is not known whether they
have 5'-terminal genome-linked proteins (VPg) or m7G RNA cap
structures. However, the biological activity of RNA transcripts from full-length
cDNA clones of the satellite RNA is unaffected by addition of a cap
(Taliansky & Robinson, 1997a).
The GRV genomic RNA sequence
(Taliansky et al., 1996)
contains four
large open reading frames (ORFs)
(Fig. 11).
ORF2 includes sequences that
encode motifs characteristic of viral RNA-dependent RNA polymerases and is
probably expressed by a -1 frameshift mechanism as a fusion with the ORF1
product. ORFs 3 and 4 almost completely overlap in different reading frames
and are probably expressed from subgenomic RNA
(Taliansky et al., 1996).
ORF4 codes for a 28 K protein which is involved in virus movement
(Ryabov et al., 1998);
its amino acid sequence has significant similarity
with those of several other viral movement proteins, especially with the 3a
protein of
cucumber mosaic virus (CMV).
The function of the ORF3 product is
not known; it has significant sequence similarity with the corresponding products
of the umbraviruses
carrot mottle mimic virus
(Gibbs et al., 1996a,
1996b)
and pea enation mosaic virus-2
(Taliansky et al., 1996),
but not with any other viral or non-viral proteins in existing databases. None of
the putative protein products of GRV RNA seems to be a particle protein.
The ten GRV satellite RNA sequences determined by Blok et al. (1994) contained from two to five short ORFs, but mutational analysis (Taliansky & Robinson,1997b) has shown that no satellite-encoded ORFs are required, either for satellite replication or for symptom induction in N. benthamiana. However, three functional untranslated RNA elements have been identified. One (designated R) is essential for RNA replication and appears to act in cis. Symptom induction in N. benthamiana and groundnut involves two further untranslated RNA elements acting in trans: (A) located in the left half, and (B) located in the right half of the satellite RNA molecule. Element A contains the determinant that is unique to the yellow blotch satellite.
Relations with Cells and Tissues
The localization of the proteins encoded by GRV ORFs 3 and 4 was studied in Nicotiana spp. by expressing the proteins as fusions with the jellyfish green fluorescent protein (GFP) from modified PVX and tobacco mosaic virus (TMV) vectors (Ryabov et al., 1998). The GFP-GRV ORF3 fusion protein was found in granules associated with membranes of the endoplasmic reticulum, and accumulated mainly in large cytoplasmic inclusion bodies lying close to the nuclei. This protein was also detected in nucleoli. The GRV ORF4-GFP fusion product was associated with the plasmodesmata of epidermal and mesophyll cells, as is found with several other viral movement proteins, including the 3a movement protein of CMV, which has sequence similarity with the GRV ORF4 protein. In addition, the GRV ORF4-GFP fusion product was detected in the cytoplasmic inclusion bodies of mesophyll cells.
Notes
Groundnuts, which came originally from South America, are cultivated worldwide
throughout the tropics and sub-tropics, but only in Africa are they affected by
rosette disease. This suggests that the causal agents of rosette are endemic to
Africa and are pathogens of some wild African plant species. However,
groundnut is the only plant in which GRV,
GRAV
or the GRV satellite have been
found occurring naturally. Experimentally, GRV and GRAV have each been
transmitted to a few other plant species but the only known hosts of both viruses,
apart from groundnut,
are Stylosanthes gracilis, S. mucronata, S. sundaica, Trifolium incarnatum, T.
repens, Gomphrena globosa and Spinacia oleracea, and none of
these is likely to act as a dry season host of Aphis craccivora. However,
since neither of the viruses is seed-borne it seems likely that at least one
species that is a host of the virus complex and of the aphid vector must exist.
There is disagreement
(Evans, 1954;
Booker, 1963;
Hildebrand et al., 1991)
about the extent to which persistence of the viruses through the dry
season may now depend on survival of infected groundnut plants
(‘groundkeepers’) in favoured areas, such as around watercourses. In regions
where this does not happen, initial infection may depend on the influx of
viruliferous aphids from other parts of Africa on prevailing winds
(Bunting, 1950;
Adams, 1967;
Rossel, 1977).
Ways of protecting groundnut crops against rosette disease include the removal
of groundkeepers, early sowing at high plant density (which reduces disease
incidence because the landing response of the aphid vector is inhibited as the
ground becomes covered by plant growth;
Booker, 1963;
A'Brook, 1964;
Farrell, 1976)
and, if affordable, the use of insecticides
(Davies, 1972).
However, the
best approach lies in the development of resistant cultivars. Resistance is
currently available in late-maturing cultivars derived from groundnut material
found in the border region between Côte d'Ivoire and Burkina Faso
(Sauger & Catharinet, 1954;
De Berchoux, 1958).
The resistance, which
does not amount to absolute immunity, is controlled by two independent
recessive genes and is effective against both chlorotic and green forms of
rosette
(Nigam & Bock, 1990;
Olorunju et al., 1992).
The resistance
is directed against GRV (and therefore the GRV satellite RNA) but is not
effective against GRAV
(Bock et al., 1990).
Recently, progress has been
made in transferring this form of resistance into early-maturing cultivars, which
are the type needed in most parts of Africa. Another form of resistance may
exist in the wild diploid species Arachis chacoense, which seems immune
to both GRV and GRAV
(Murant et al., 1991):
a high degree of resistance
to rosette was found in a hybrid derivative from an interspecific cross of A.
hypogaea x A. chacoense
(Moss et al., 1993).
A possibility for the future is the deployment of transgenic forms of resistance.
Because of the relative difficulty in transforming groundnut, the use of constructs
directed against GRV has been evaluated in Nicotiana benthamiana
(Taliansky et al., 1998).
Resistance to GRV infection was detected in
plants transformed with constructs derived from a mild variant of the satellite
RNA. However, this strategy has not yet been tested for protection against
rosette disease in groundnut.
Resistance in groundnut to Aphis craccivora was reported by Padgham et al. (1990) and this could prove a useful additional approach, especially because the aphid is an important pest in its own right.
Acknowledgements
All photographs copyright Scottish Crop Research Institute.
Figures

Groundnut plant infected with chlorotic rosette in Malawi, showing typical leaf symptoms.

Foreground, groundnut plant infected with chlorotic rosette in Malawi showing extreme stunting compared with the apparently healthy plant behind it.

Experimentally infected groundnut plant showing symptoms of chlorotic rosette.

Experimentally infected groundnut plant showing symptoms of mosaic rosette.

Experimentally infected groundnut plant showing symptoms of green rosette.

Local lesions induced by GRV in Chenopodium amaranticolor.

Nicotiana benthamiana plant infected with a GRV isolate carrying a normal satellite RNA, showing local spots and rings on inoculated leaf (left of picture) and necrotic patterns on the first systemically infected leaf (bottom of picture).

Nicotiana benthamiana plant infected with a GRV isolate carrying the yellow blotch satellite RNA, showing brilliant yellow systemic symptoms.

Nicotiana clevelandii plant infected with GRV, showing systemic mottle, crinkling and distortion of apical leaves.

Electrophoresis in 7% polyacrylamide gel of dsRNA preparations from N. benthamiana inoculated with (lane A) a satellite-containing isolate of GRV, (lane B), an experimentally produced satellite-free isolate. Arrows 1-3 indicate dsRNA-1, dsRNA-2 and dsRNA-3, respectively. Lane C, dsRNA of rice dwarf virus used as a size standard.
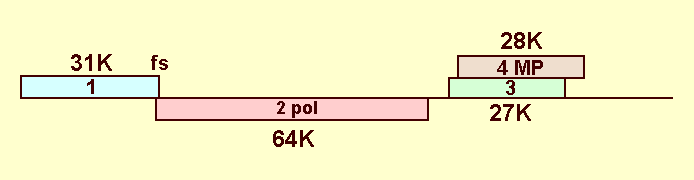
Diagram showing the arrangement of the ORFs in GRV RNA. The continuous horizontal line represents the RNA and the numbered coloured blocks the correspondingly numbered ORFs. The mol. wt of the predicted translation product is shown adjacent to each ORF. The ORF encoding a probable RNA-dependent RNA polymerase is marked ‘pol’, and that encoding a probable cell-to-cell movement protein is marked ‘MP’. The position of the frame-shift event is marked ‘fs’.
References list for DPV: Groundnut rosette virus (355)
- A’Brook, Ann. appl. Biol. 54: 199, 1964.
- Adams, Rhodesia, Zambia Malawi J. agric. Res. 4: 125, 1966.
- Adams, Rhodesia, Zambia Malawi J. agric. Res. 5: 145, 1967.
- Anon., Ann. Progr. Rep., SADC/ICRISAT Groundnut Project, 1996, Chitedze Research Station, Malawi, 1996.
- Blok, Ziegler, Robinson & Murant, Virology 202: 25, 1994.
- Blok, Ziegler, Scott, Dangora, Robinson & Murant, Ann. appl. Biol. 127: 321, 1995.
- Bock, Murant, & Rajeshwari, Ann. appl. Biol. 117: 379, 1990.
- Booker, Ann. appl. Biol. 52: 125, 1963.
- Breyel, Casper, Ansa, Kuhn, Misari & Demski, J. Phytopath. 121: 118, 1988.
- Bunting, Ann. appl. Biol. 37: 699, 1950.
- Davies, Bull. ent. Res. 62: 169, 1972.
- De Berchoux, Oléagineux 13: 237, 1958.
- Demler, Rucker, de Zoeten, Ziegler, Robinson & Murant, J. gen. Virol. 77: 2847, 1996.
- Dodds & Bar-Joseph, Phytopathology 73: 419, 1983.
- Dubern, Phytopath. Z. 99: 318, 1980.
- Evans, Ann. appl. Biol. 41: 189, 1954.
- Farrell, Bull. ent. Res. 66: 317, 1976.
- Gibbs, Cooper & Waterhouse, Virology 224: 310, 1996a.
- Gibbs, Ziegler, Robinson, Waterhouse & Cooper, Molec. Pl. Path. On-Line 1996/1111 gibbs, 1996b.
- Hayes, Trop. Agric., Trinidad 9: 211, 1932.
- Hildebrand, Bock & Nigam, p.8, in: Groundnut Virus Diseases in Africa, Proc. 4th Meeting Consult. Gp Collab. Res. Groundnut Rosette Virus Disease, Montpellier, France 1990, ICRISAT, 1991.
- Hull & Adams, Ann. appl. Biol. 62: 139, 1968.
- Kumar, Murant & Robinson, Ann. appl. Biol. 118: 555, 1991.
- Mayo & D’Arcy, in: The Luteoviridae, Smith & Barker, eds, Wallingford: CAB International, in press, 1998.
- Misari, Abraham, Demski, Ansa, Kuhn, Casper & Breyel, Pl. Dis. 72: 250, 1988.
- Moss, Singh, Subrahmanyam, Hildebrand & Murant, Int. Arachis Newsl. No. 13, 22, 1993.
- Murant, J. gen. Virol. 71: 2163, 1990a.
- Murant, Rep. Scott. Crop Res. Inst., 1989, 138, 1990b.
- Murant & Kumar, Ann. appl. Biol. 117: 85, 1990.
- Murant, Rajeshwari, Robinson & Raschke, J. gen. Virol. 69: 1479, 1988.
- Murant, Kumar & Robinson, p.7, in: Groundnut Virus Diseases in Africa, Proc. 4th Meeting Consult. Gp Collab. Res. Groundnut Rosette Virus Disease, Montpellier, France 1990, ICRISAT, 1991.
- Murant, Robinson, Blok, Scott, Torrance & Farmer, Ann. Rep. Scott. Crop Res. Inst., 1992, 85, 1993.
- Murant, Robinson & Gibbs, pp. 388-391, in: Virus Taxonomy - Classification and Nomenclature of Viruses. 6th Rep. Int. Comm. Taxonomy of Viruses, Murphy et al., eds, Vienna: Springer-Verlag, 1995.
- Nigam & Bock, Ann. appl. Biol. 117: 553, 1990.
- Okusanya & Watson, Ann. appl. Biol. 58: 377, 1966.
- Olorunju, Kuhn, Demski, Misari & Ansa, Pl. Dis. 76: 95, 1992.
- Padgham, Kimmins & Ranga Rao, Ann. appl. Biol. 117: 285, 1990.
- Rajeshwari & Murant, Ann. appl. Biol. 112: 403, 1988.
- Reddy, Murant, Duncan, Ansa, Demski & Kuhn, Ann. appl. Biol. 107: 57, 1985a.
- Reddy, Murant, Raschke, Mayo & Ansa, Ann. appl. Biol. 107: 65, 1985b.
- Robinson & Taliansky, Ann. Rep. Scott. Crop Res. Inst., 1996/7, 170, 1997.
- Rossel, Trop. Grain Legume Bull. 8: 41, 1977.
- Ryabov, Oparka, Santa Cruz, Robinson & Taliansky, Virology 242: 303, 1998.
- Sauger & Catharinet, L'agronomie tropicale 9: 28, 1954.
- Smartt, Emp. J. exp. Agric. 29: 79, 1961.
- Storey & Bottomley, Ann. appl. Biol. 15: 26, 1928.
- Storey & Ryland, Ann. appl. Biol. 43: 423, 1955.
- Storey & Ryland, Ann. appl. Biol. 45: 318, 1957.
- Subrahmanyam & Chiyembekeza, Int. Arachis Newsl. No.15, 22, 1995.
- Subrahmanyam & Mamba, Int. Arachis Newsl. No.13, 7, 1993.
- Taliansky & Robinson, Virology 230: 228, 1997a.
- Taliansky & Robinson, J. gen. Virol. 78: 1277, 1997b.
- Taliansky, Robinson & Murant, J. gen. Virol. 77, 2335, 1996.
- Taliansky, Ryabov & Robinson, Mol. Pl. Microbe Interact. 11: 367, 1998.
- Watson & Okusanya, Ann. appl. Biol. 60: 199, 1967.
- Yaycock, Rossel & Harkness, Samaru Conference Paper No. 9, Institute of Agricultural Research, Samaru, Nigeria, 1976.
- Zimmermann, Der Pflanzer 3: 129, 1907.
- Olorunju, Kuhn, Ansa, Misari & Demski, Peanut Sci. 22: 56, 1995.