Details of DPV and References
DPV NO: 37 October 1970
Family: Potyviridae
Genus: Potyvirus
Species: Potato virus Y | Acronym: PVY
There is a more recent description of this virus: DPV 242
Potato virus Y
S. Delgado-Sanchez Instituto Nacional de Investigaciones Agricolas, Apdo. Postal No. 6-882, Mexico 6, D. F. Mexico
R. G. Grogan Dept. of Plant Pathology, Univ. of California, Davis, California 95616, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Smith (1931).
Selected synonyms
- Potato acropetal necrosis virus (Rev. appl. Mycol. 10: 745)
- Tobacco vein-banding virus (Rev. appl. Mycol. 10: 60)
- Tobacco veinal necrosis virus (Rev. appl. Mycol. 31: 201)
- Solanum virus 2 (Rev. appl. Mycol. 17: 52)
- Marmor upsilon (Rev. appl. Mycol. 28: 514)
- Tobacco vein-banding virus (Rev. appl. Mycol. 10: 60)
-
A virus with flexuous helically constructed particles, 730 x 11 nm, sap-transmissible to a narrow range of hosts and transmitted by several species of aphids in the non-persistent manner. It causes important economic diseases of solanaceous plants and has a world-wide distribution.
Main Diseases
In potato it usually causes ‘leaf-drop streak’ or necrosis along the veins of the underside of leaflets in the first year of infection (Fig. 3, Fig. 4); in combination with potato virus X it causes the disease called ‘rugose mosaic’ in the USA, one of the most destructive diseases of potato. In Britain, the term, rugose mosaic, is used to refer to disease caused by potato virus Y in the second and subsequent years of infection. In tobacco, pepper, and tomato the virus causes mild to severe mottling depending on the strain (Smith, 1957).
Geographical Distribution
World-wide.
Host Range and Symptomatology
Reported to infect at least 60 plant spp., mostly in the Solanaceae, but also infects members of the Chenopodiaceae and Leguminosae (Thornberry, 1966).
-
Diagnostic species
- Capsicum frutescens
cv. Tabasco (pepper). Mild to severe mottle; no wilting like that caused by tobacco etch virus. - Datura stramonium. Immune to infection by all tested strains of
potato virus Y, but susceptible
to tobacco etch virus; useful for separating potato virus Y from tobacco etch virus.
- Nicotiana glutinosa. Mild to severe mottling depending on the strain (Fig. 1).
- N. tabacum (tobacco). Most strains produce vein clearing followed by mottling; an exception is the necrotic strain. Combined infection with potato viruses X and Y produces a synergistic ‘spot necrosis’ reaction which has been used to detect the presence of potato virus Y (Darby, Larson & Walker, 1951).
- Solanum tuberosum cv. Saco (potato). Some strains produce local necrotic lesions; useful for separating potato virus Y from potato virus X to which Saco is virtually immune (Benson & Hooker, 1960).
- Nicotiana glutinosa. Mild to severe mottling depending on the strain (Fig. 1).
-
Propagation species
- N. glutinosa
may be used to maintain the virus. N. tabacum cv. Wisconsin Havana 425 is a good source for virus purification.Assay species
- Local lesion hosts include Chenopodium amaranticolor, C. quinoa (Fig. 5), several species of Lycium, Nicandra physalodes, Nicotiana repanda, N. rustica, Physalis floridana, Solanum demissum ‘Y’ (Fig. 2), S. demissum x S. tuberosum ‘A-6’, and S. tuberosum cvs. Saco and U.S.D.A. seedling 41956.
Strains
Several strains can be distinguished on the basis of severity of systemic symptoms
in tobacco and
other hosts. The following seem the most important:
Common strains: Differentiated by the severity of symptoms in
N. glutinosa, P. floridana
and potato. Found world-wide
(Kahn & Monroe, 1963).
The necrotic or tobacco veinal necrosis strain
(Nobrega & Silberschmidt, 1944)
produces
severe veinal necrosis in tobacco. Source South America.
Potato virus C (Bawden, 1936) differs from others in not being transmitted by Myzus persicae, which is an efficient vector of PVY. Source Australia and England.
Transmission by Vectors
Transmitted in the non-persistent manner by several aphid spp. Myzus persicae is probably the most efficient vector; others are Myzus ornatus, Macrosiphum euphorbiae, Aulacorthum circumflexum, Aphis nasturtii and Aphis gossypii (Kennedy, Day & Eastop, 1962). Young upper leaves of infected potato plants are better sources of virus for aphids than are middle or lower leaves (Bagnall & Bradley, 1958). Adult or later instar apterous aphids have usually been used in transmission experiments and a pre-acquisition starvation period of 2-7 h aids transmission. The optimum acquisition feeding period is from 30 sec to 5 min a period of 30-60 sec to 24 h is satisfactory for inoculation (Bradley, 1953; Easton, Larson & Hougas, 1958).
Transmission through Seed
Not reported.
Transmission by Dodder
Not reported.
Serology
Strongly immunogenic. Emulsification of antigen with Freund’s adjuvant before injecting it intramuscularly into rabbits increased both the titre of the antiserum obtained and the period of maximum immune response (Wetter, 1960). Microprecipitin tests can be done using clarified extracts of infected tissue. The formation of non-specific precipitates can be avoided by buffering the saline solution with 0.05 M Tris-HCl. Serological precipitates in mixed liquids are of the flocculent (flagellar) type.
Relationships
Precipitin tests showed that the common, C and necrotic strains are serologically related (Bawden & Kassanis, 1951), but differences were noted between the homologous and heterologous reactions, particularly between the necrotic and the common strains (Bartels, 1957). Using high titred antisera, PVY was shown to be distantly related to at least 16 other filamentous viruses, and these comprise the potato virus Y group (Brandes & Bercks, 1965).
In cross-protection tests, several mild isolates protected against more virulent isolates in potato, but not in tobacco (Darby et al., 1951).
Stability in Sap
Infectivity is lost after heating sap from infected tobacco for 10 min at 55-60°C; dilution end-points range from 10-2 to 10-3 for most isolates; inactivated in vitro within 48-72 h.
Purification
Several procedures have been described, but aggregation of the particles presents difficulties (Van Regenmortel, 1964; Venekamp & Mosch, 1964). A procedure for obtaining essentially unaggregated preparations has been described (Delgado-Sanchez & Grogan, 1966).
Particle Structure
Virus particles are flexuous filaments about 11 nm wide and helically constructed, with a pitch of about 3.3 nm (Varma et al., 1968). The normal length of particles in purified virus preparations is about 684 nm, compared with about 730 nm for particles in tissue-dip preparations (Fig. 6).
Particle Composition
Not known.
Relations with Cells and Tissues
Pin-wheel inclusions (Fig. 8) occur in systemically infected tissue (Edwardson, 1966). They are composed of pin-wheel plates, which are easily seen in leaf-dip preparations (Fig. 7) and have characteristic striations with a spacing of c. 5 nm.
Notes
In some Solanum species the virus may be confused with tobacco etch and henbane mosaic viruses, which produce somewhat similar symptoms and have similar host ranges. Tobacco etch virus, however, infects Datura stramonium and produces necrotic wilting of Tabasco pepper and etching of the leaves of tobacco. Henbane mosaic virus occurs less frequently in the common hosts of potato Y and tobacco etch viruses; it can infect D. stramonium but not Tabasco pepper.
Figures

Systemic mottle in Nicotiana glutinosa.

Necrotic local lesions in inoculated leaves of Solanum demissum ‘Y’.

Symptoms of ‘leaf-drop streak’ in potato.

Systemic necrotic symptoms in potato leaf.

Chlorotic local lesions in inoculated leaves of Chenopodium quinoa.

Flexuous elongated virus particles in uranyl formate. Bar represents 200 nm.

Pinwheel plate in leaf-dip preparation mounted in phosphotungstate. Bar represents 100 nm.
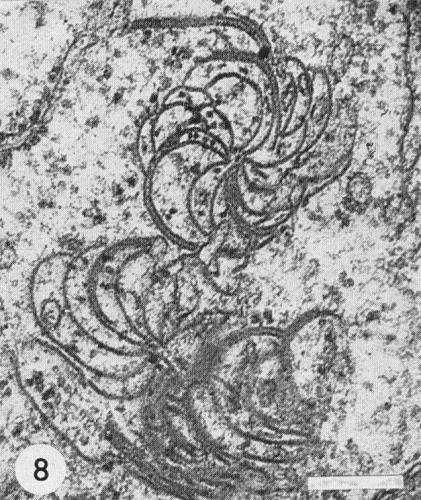
Pinwheel inclusion bodies typically produced in tissues infected with potato virus Y and other viruses in the potato virus Y group. Bar represents 200 nm.
References list for DPV: Potato virus Y (37)
- Bagnall & Bradley, Phytopathology 48: 121, 1958.
- Bartels, Proc. 3rd Conf. on Potato Virus Diseases, Lisse-Wageningen: 13, 1957.
- Bawden, Ann. appl. Biol. 23: 487, 1936.
- Bawden & Kassanis, Ann. appl. Biol. 38: 402, 1951.
- Benson & Hooker, Phytopathology 50: 231, 1960.
- Bradley, Nature, Lond. 171: 755, 1953.
- Brandes & Bercks, Adv. Virus Res. 11: 1, 1965.
- Darby, Larson & Walker, Res. Bull. Univ. Wisc. 177, 32 pp., 1951.
- Delgado-Sanchez & Grogan, Phytopathology 56: 1397, 1966.
- Easton, Larson & Hougas, Res. Bull. Univ. Wisc. 205, 32 pp., 1958.
- Edwardson, Am. J. Bot. 53: 359, 1966.
- Kahn & Monroe, Phytopathology 53: 1356, 1963.
- Kennedy, Day & Eastop, A conspectus of aphids as vectors of plant viruses, London, Commonwealth Institute of Entomology, 1962.
- Nobrega & Silberschmidt, Arquiv. Inst. Biol. S. Paulo 15: 307, 1944.
- Smith, Proc. R. Soc., B 109: 251, 1931.
- Smith, A Text Book of Plant Virus Diseases, Churchill, London, 1957.
- Thornberry, Index of Plant Virus Diseases, Agric. Handb. U.S.D.A., 307, 285, 1966.
- Van Regenmortel, Virology 23: 495, 1964.
- Varma, Gibbs, Woods & Finch, J. gen. Virol. 2: 107, 1968.
- Venekamp & Mosch, Virology 23: 394, 1964.
- Wetter, Arch. Mikrobiol. 37: 278, 1960.