Details of DPV and References
DPV NO: 371 March 2000
Family: Tombusviridae
Genus: Dianthovirus
Species: Carnation ringspot virus | Acronym: CRSV
This is a revised version of DPV 308
Carnation ringspot virus
Steven A. Lommel Department of Plant Pathology, North Carolina State University, Box 7616, Raleigh, NC 27695-7616, USA.
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Kassanis (1955).
Synonym Anjermozaiek virus (Noordam et al., 1951).
A virus with icosahedral isometric particles approximately 34 nm in diameter composed of 180 copies of a Mr 38,000 capsid protein and two genomic ssRNA species of 3.8 and 1.4 kb. The virus was once cosmopolitan in carnations, but is now only infrequently detected due to rigorous detection and sanitation. It also infects grapevine and a number of orchard crops in Central and Eastern Europe. It is readily mechanically transmitted and is also transmitted through the soil without the aid of a biological vector.
Main Diseases
Infection in carnations results in diagnostic ringspots, mottling, and leaf and flower distortions (Fig. 1, Fig. 2, Fig.3; Hollings & Stone, 1970; Kowalska, 1972; Lovisolo & Lisa, 1978). The disease symptoms are enhanced when carnations are co-infected with carnation mottle virus (CarMV; Family Tombusviridae, Genus Carmovirus) (Kemp, 1964; Hollings & Stone, 1965). In severe infections or in susceptible carnation cultivars, leaf tip necrosis can also be observed. In general, CRSV infections do not kill the host plants, but necrosis and other symptoms can become more severe at sustained temperatures between 15 and 20°C. Symptoms are greatly reduced at temperatures above 20°C. Symptoms in stone fruits and other orchard crops may be mild and difficult to discern (Hiruki, 1987). CRSV causes a disease termed "stony pit" in pear (Richter et al., 1978) and decline diseases in sour cherry and apple (Kleinhempel et al., 1980).
Geographical Distribution
Identified wherever carnations are grown on a large scale (Lommel et al., 1983b). Also established in stone fruit and grape orchards in Central and Eastern Europe (Richter et al., 1978; Fritzsche et al., 1979; Kleinhempel et al., 1980; Kegler et al., 1983).
Host Range and Symptomatology
Transmitted experimentally to over 133 species in 25 families (Hollings & Stone, 1970; Kleinhempel et al., 1980). In nature, infects carnations (Dianthus caryophyllus) and a wide range of orchard trees (including plum, pear, apple, sour cherry, and sweet cherry), and grapevines (Richter et al., 1978; Fritzsche et al., 1979; Kleinhempel et al., 1980; Kegler et al., 1983). Has also been identified in several weed species, particularly Stellaria media growing within infected fruit orchards (Rudel et al., 1977; Fritzsche et al., 1979; Kegler et al., 1983). The experimental host range is much broader than that found in nature (Hollings & Stone, 1970). Easily mechanically transmissible to a wide range of herbaceous species, and can systemically infect a number of members of the Solanaceae, Leguminosae, Cucurbitaceae and Compositae. Infects an even larger number of plants non-systemically (Hollings & Stone, 1970). On experimental systemic hosts, causes concentric ringspots with necrotic centres on the inoculated leaves and mosaics, necrotic flecks and often veinal necrosis on the systemically infected leaves (Hollings & Stone, 1970).
- Diagnostic species
Chenopodium amaranticolor and C. quinoa. Local necrotic lesions in 2-4 days (Fig. 4); usually not systemic.
Dianthus barbatus (Sweet William). Inoculated leaves exhibit flecks, rings and ringspots after 4-7 days (Fig. 5) followed by systemic chlorotic and semi-necrotic rings and flecks. Not all clones exhibit obvious symptoms or support large concentrations of the virus.
Gomphrena globosa. Local necrotic rings develop in 2-4 days after inoculation followed by systemic flecking, mottle and distortion.
Phaseolus vulgaris (French bean). Local chlorotic dots in 4-5 days, becoming white and necrotic; irregular systemic spotting and necrotic veinal flecks, later growth symptomless though infected.
Tetragonia expansa. Local white necrotic dots in 2-3 days, sometimes followed by systemic necrotic flecks.
Vigna unguiculata ssp. sinensis (cowpea). Local necrotic lesions in 2-4 days (Fig. 6), followed by systemic mottling, necrotic flecking, rugosity and rolling of leaf laminae (Weintraub et al., 1975).
- Propagation species
Dianthus barbatus is an acceptable host for maintaining cultures. Nicotiana clevelandii, and to a lesser extent Phaseolus vulgaris and Vigna unguiculata, are good hosts from which to purify the virus.
- Assay species
Chenopodium amaranticolor, C. quinoa and Vigna unguiculata ssp. sinensis are useful local lesion hosts.
Strains
Strains based on differential host reactions have not been reported. Several strains are reported based on distinctive aggregation properties. The A strain assembles into clusters of 12 particles assuming an icosahedral symmetry. This 12-particle unit can form higher order aggregates. N and R strains form alternative two-dimensional net-like aggregates (Tremaine et al., 1976).
Transmission by Vectors
Not transmitted by insects or soil-inhabiting fungi (Hiruki, 1987). Confusion exists as to whether transmission occurs from plant to plant by nematodes. Early reports suggested transmission by several nematode species including Longidorus elongatus, L. macrosoma and Xiphinema diversicaudatum (Fritzsche & Schmelzer, 1967; Kegler et al., 1977). More recent reports indicate that virus particles are released directly from infected roots into the soil. Plant-to-plant transmission then occurs passively through the soil in the absence of a biological vector (Kegler & Kegler, 1981; Brown & Trudgill, 1984). Virus particles have been detected in drainage canals downstream from infected orchards (Koenig et al., 1988, 1989). Presumably, infection occurs through microscopic root wounds. Nematode and soil fungus colonization of roots increases the possibility of soil transmission by generating virus entry sites, but transmission is not absolutely dependent on this. In nature, the virus is transmitted from plant to plant by physical contact or by contaminated soil or by vegetative propagation (Lommel et al., 1983b).
Transmission through Seed
There are no published reports of transmission through the seeds of infected plants.
Serology
CRSV is a moderate to strong immunogen, and a number of high quality polyclonal antisera are available (Hiruki, 1987). Antisera with gel diffusion titres up to 1/1024 have been obtained. Agar double-diffusion (Kowalska, 1972) and several variations of ELISA (Lommel et al., 1982) are effective in identifying the virus. Antibodies prepared by injecting rabbits with particles do not react in gel diffusion tests with dissociated protein subunits but do react with these antigens in indirect ELISA (Tremaine & Dodds, 1985). Strain differences have been detected in gel diffusion tests (Tremaine et al., 1976; 1984).
Relationships
CRSV is the type member of the Dianthovirus genus which contains two other members, red clover necrotic mosaic virus (RCNMV) and sweet clover necrotic mosaic virus (SCNMV) (Lommel, 1995). CRSV can potentially be confused with carnation mottle virus (CarMV), which is widely distributed in commercial carnations (Lommel et al., 1983a) and consequently occurs frequently together with CRSV in mixed infections (Kemp, 1964; Hollings & Stone, 1965). CRSV and CarMV are indistinguishable when examined in the electron microscope with negative stain, yet are serologically distinct and are readily distinguished by differential symptoms on Gomphrena globosa and Dianthus barbatus (see Description No. 7). CarMV is the type member of the Carmovirus genus. CRSV is similar in name only to carnation Italian ringspot virus, which is in the genus Tombusvirus (Russo et al., 1987). Like the dianthoviruses, the carmoviruses and tombusviruses belong to the family Tombusviridae (Lommel, 1995).
Stability in Sap
In Dianthus barbatus sap, the thermal inactivation point (10 min) is about 80°C although much infectivity is lost above 60°C. Dilution end-point is 10-5 and infectivity is retained at 20°C for 50-60 days. In lyophilized Nicotiana clevelandii sap the virus retained infectivity at room temperature, under vacuum, for over 6 years (Hollings & Stone, 1970).
Purification
Easily purified from infected Nicotiana clevelandii or cowpea plants. Fresh tissue can yield 150 mg virus/kg; frozen tissue yields significantly less virus.
Method 1 (Tremaine et al., 1976). Extract 100 g infected tissue 3 weeks after inoculation in 200 ml 0.2 M sodium acetate buffer (pH 5.0) containing 20 mM sodium diethyl dithiocarbamate and 0.1% 2-mercaptoethanol. Adjust the extract to pH 5.0 with 10% acetic acid, and leave at 4°C for 4 h. Clarify by low speed centrifugation and precipitate the virus particles by adding polyethylene glycol (mol. wt 6000) to 8.0% (w/v). Suspend the pellet from low speed centrifugation in 0.1 M sodium acetate buffer (pH 5.0) and give the preparation one cycle of differential centrifugation (20 min at 12,000 g; 90 min at 95,000 g). Further purify by repeated cycles of differential centrifugation and rate zonal sucrose density gradient centrifugation.
Method 2 (Hollings & Stone, 1965). Must be used for strains that precipitate at pH 5.0. Extract each 100 g tissue in 125 ml 50 mM phosphate buffer (pH 7.6) containing 0.1% thioglycollic acid. Clarify by stirring with 8.5% butan-1-ol overnight at 2°C. The virus particles are then concentrated and purified by differential centrifugation.
Properties of Particles
The particles swell at pH >7.0. Swelling is prevented or reversed by divalent cations. The particles dissociate in 0.1 M tris-HCl buffer, pH 7.5, containing 10 mM EDTA and 1 M NaCl. Stable particles can be reconstituted by dialysis in 0.1 M tris-HCl , pH 7.0 with divalent cations (Tremaine & Dodds, 1985). The particles are stabilized by pH-dependent protein-protein interactions and by RNA- protein interactions. Strain A particles can form aggregates of six virus particles and linked aggregates (Fig. 7). R and N strain particles aggregate in a temperature-reversible manner but the transition occurs at lower temperatures with particles of the R strain than with those of the N strain. N strain particles are readily dissociated at room temperature in low concentrations of SDS at pH 7.0. A and R strain particles are more stable than N strain particles (Tremaine & Ronald, 1976; Ronald & Tremaine, 1976; Tremaine et al., 1976, 1983, 1984).
Sedimentation coefficient (s20,w): N strain 133 S at pH 5.0, 125 S at pH 7.5 (Tremaine & Dodds, 1985).
Particle weight: 8.526 x 106 daltons based on calculations from the complete nucleotide sequence and assuming that the shell contains 180 copies of the capsid protein (Kendall & Lommel, 1992; Ryabov et al., 1994).
Electrophoretic mobility: in free boundary electrophoresis the virus particles are isoelectric at pH 4.5; at pH 6.0 to 8.0 the virus boundary moves to the anode as an increasingly broad Schlieren peak which continues to broaden when the polarity is reversed (Tremaine & Dodds, 1985).
Diffusion coefficient (D20,w x 10-7 cm2/sec): 1.48 (Kalmakoff & Tremaine, 1967).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 6.46 (Kalmakoff & Tremaine, 1967).
A260/A280: 1.67 corrected for light-scattering (Tremaine & Dodds, 1985).
Buoyant density in CsCl: 1.366 g/cm3. When limited proteolysis of virus occurs, additional species with buoyant densities of 1.369 and 1.374 are present.
Particle Structure
The particles are 34 nm icosahedra with a T=3 symmetry (Fig. 8; Kalmakoff & Tremaine, 1967). The particles have distinctively granular surfaces as also found in species of Tombusvirus and Carmovirus (Lommel, 1995). The protein shell, composed of 180 copies of the 38 kd capsid protein, packages the 3.8 kb and 1.4 kb genomic ssRNAs (Kendall & Lommel, 1992; Ryabov et al., 1994; Lommel, 1995). The mol. wt of a virion, assuming that each particle possesses one copy each of RNA-1 and RNA-2 is 8.5 MDa (Kuhne et al., 1985; Tremaine & Dodds, 1985). However, it is not certain that this is the packaging arrangement (see PARTICLE COMPOSITION).
Particle Composition
Nucleic acid. ssRNA, constituting 20% of the particle weight. The genome is composed of two species of RNA termed RNA-1 and RNA-2 (Fig. 9). It is not known how the RNAs are packaged into virions (Hamilton & Tremaine, 1996). Particle preparations yield 2-3 times as many molecules of RNA-2 as of RNA-1, so that some particles must contain RNA-2 alone. Some evidence suggests that each particle contains one copy of each RNA, whereas other evidence suggests that there are two particle classes, one containing one molecule of RNA-1 and the other containing three molecules of RNA-2. RNA-1, 3840 nt (accession no. L18870; Ryabov et al., 1994) with a mol. wt of 1.246 x 106, is composed of 1037 A, 903 C, 972 G and 928 U residues for molar percentages of 27.01%, 23.52%, 25.31%, and 24.17%, respectively. RNA-2, 1403 nt (accession no. M88589; Kendall & Lommel, 1992) with a mol. wt of 0.455 x 106, is composed of 380 A, 364 U, 318 C, and 340 G residues for molar percentages of 27.1%, 25.96%, 22.68%, and 24.25%, respectively.
Protein. Capsid protein can be isolated by first swelling the virus at 5-10 mg/ml in 0.1 M tris-HCl, 10 mM EDTA, pH 7.5 to 8.0, for at least 1 h at 0°C, and then adding an equal volume of 2.0 M NaCl and subjecting the mixture to rate zonal density gradient centrifugation in a 5-35% sucrose density gradient in 0.1 M tris-HCl buffer, pH 7.0, containing 1.0 M NaCl (Tremaine & Dodds, 1985). Virus particles contain a single polypeptide species comprising 80% of the particle weight. The polypeptide has a Mr of 37,900 but limited proteolysis may occur in stored preparations to yield polypeptides of Mr 36,000 and 34,000. The polypeptide is composed of 345 amino acid residues.
Genome Properties
Based on research conducted with RCNMV, another virus in the same genus, the two genomic RNAs of CRSV are probably infectious as naked nucleic acids. CRSV RNA-1 replicates in plant protoplasts in the absence of RNA-2 (Paje-Manalo & Lommel, 1989), but both RNA-1 and RNA-2 are required for infectivity on plants. The 3'-proximal capsid protein ORF located on RNA-1 is expressed in vivo from a ~1.5 kb subgenomic RNA (Fig. 10; Zavriev et al., 1996). Three dsRNAs accumulate in infected cells corresponding to the two genomic RNAs and the subgenomic RNA (Fig. 9). A sequence element within RNA-2 is required to trans-activate the synthesis of subgenomic RNA from RNA-1 (Sit et al., 1998). By analogy with RCNMV, each CRSV RNA is presumed to be capped at the 5' terminus with m7GpppA and is not polyadenylated at the 3'-terminus. Only the 5'-terminal 2 nt and 20 out of the 3'-terminal 26 nt are identical between RNA-1 and RNA-2. The near identical 3'-terminal 26 nt are capable of forming similar stem-loop structures.
RNA-1 encodes two ORFs. The first ORF (p88) initiates at the first methionine codon, 58 nt from the 5' terminus (Fig. 10). The p88 ORF is capable of encoding a 769 amino acid Mr 87,776 polypeptide. This ORF is interrupted by a retrovirus-like -1 ribosomal frameshifting element (Kim & Lommel, 1998) yielding a pre-readthrough 236 amino acid Mr 27,158 polypeptide (p27) that is identical to the first 236 amino acids of the 88 kd protein (Kim & Lommel, 1994). The 3'-most ORF encodes the 345 amino acid Mr 37, 921 capsid protein (p38). This ORF is followed by a 436 nt 3'-terminal non-coding sequence. RNA-2 contains a single ORF encoding a 303 amino acid Mr 33,730 movement protein (p34) (Kendall & Lommel, 1992). The ORF initiates at the first start codon at nt 47 and is followed by a 3'-terminal 441 nt non-coding region (Fig. 10). RNA-1 directs the synthesis of 27, 57, 88 and 37 kd polypeptides in vitro. The 57 kd polypeptide has not been observed in vivo and the independent production of this protein may well be an in vitro translation artifact (Kim & Lommel, 1994). The RNA-1-encoded 88 kd polypeptide contains the conserved Glycine-Aspartate-Aspartate ("GDD") motif present in all RNA-dependent RNA polymerases (Ryabov et al., 1994). In addition, extensive amino acid sequence similarity exists between the 27 kd and 88 kd polypeptides and the replicases encoded by all of the species within the Tombusviridae. The capsid protein is not necessary for cell-to-cell movement, but is required for rapid systemic infection through the vascular tissue. The RNA-2 encoded 34 kd movement protein is essential for cell-to-cell and long distance movement.
Relations with Cells and Tissues
The virus is found in stems, leaves, flowers, and roots as well as in some apical meristems. The virus titre is often higher in the roots than in the foliar portion of the plant. Infected cells contain large amorphous inclusions, which can be readily observed with a light microscope after staining (Fig. 11). Electron microscopic studies show that these inclusions are composed of large aggregates of virus particles within the cytoplasm. Inclusions form as the intracellular virus titre increases and become readily observable when the virus titre reaches a maximum, approximately 1 week after inoculation (Weintraub et al., 1975). Infections in cowpea result in crystalline arrays composed of virions in the cytoplasm. The nuclei also contain aggregates of virus particles, in addition to tubular inclusions (Fig. 12). In carnation, spherical inclusion bodies are also observed. Inflated and electron transparent mitochondria, as well as clusters of proliferated endoplasmic reticulum with dilated cisternae are apparent in cells with mature infections (Weintraub et al., 1975). Particles are commonly observed scattered throughout the cytoplasm of mesophyll cells, with necrotic cells having massive aggregates of virions.
Ecology and Control
An established and persistent disease problem in commercial carnations. The virus has diminished as a major problem in recent years as a result of vigilant and effective control measures (Lommel et al., 1983a). Infected nuclear stock plants, from which production cuttings are derived, serve as sources of virus inoculum in commercial glasshouses. Roguing, selection, indexing, and meristem tip culture are all effectively employed to ensure that these stock plants are free of the virus (Rybalko & Kharuta, 1978; Koev et al., 1983). Spread in carnations occurs as a result of careless propagation and leaf and root contact. In orchard crops, it has been observed that nematode infestation increases incidence (Kegler & Kegler, 1981). Cultural practices and chemicals that control nematode populations reduce the spread of the virus in orchard crops.
The most effective control is by sanitation during plant propagation. In vegetatively propagated hosts such as carnation and fruit trees, eradication of virus in plant shoots or meristems by chemo- and thermotherapy is routinely accomplished to produce certified virus free nuclear stocks which are then commercially propagated under a strict sanitation regime (Rybalko & Kharuta, 1978). More recently, antiviral agents, usually based on nucleotide analogs such as 5-fluoro- uracil, are added to the media during meristem tip culture. Production from virus-free nuclear stocks coupled with constant monitoring for CRSV infection by ELISA has ensured the production of CRSV-free commercial carnations (van Ruiten, 1987). The nearly world-wide establishment of a virus-free certified carnation propagation program has ensured that virus has not returned in epidemic forms (Ebbels, 1979).
Notes
The symptoms caused by CRSV in carnation cannot readily be confused with those caused by any other virus.
Acknowledgements
I would like to acknowledge M. Hollings and O. Stone, who prepared the first CMI/AAB Description of this virus (No. 21), and J. H. Tremaine and J. A. Dodds who prepared the first revision (No. 308). Text and figures from Descriptions No. 21 and 308 were used in part to prepare this revised description. Figs 1, 2, 4 and 6, courtesy M. Hollings & O. Stone; Fig. 7, courtesy of J. H. Tremaine; Fig. 9, courtesy of J.A. Dodds, Figs 11 and 12, courtesy of M. Weintraub.
Figures

Carnation var. Joker, (left) healthy plant, (right) infected plant showing typical symptoms of infection with CRSV (stunting and shortened internodes).
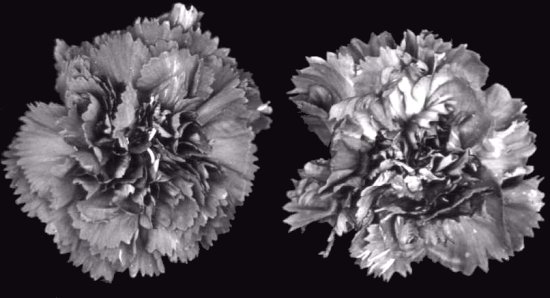
Carnation var. Joker, (left) healthy flower, (right) flower infected with CRSV.

Systemic symptoms on Dianthus caryophyllus leaves. The leaf at the top is healthy and the other three leaves are exhibiting typical symptoms including tip necrosis.

Local lesions on Chenopodium amaranticolor.

Symptoms on inoculated leaf of Dianthus barbatus.

Local lesions on Vigna unguiculata ssp. sinensis (cowpea).

Electron micrographs of particles of strain A negatively stained with 1% uranyl acetate. Bar represent 100 nm. (a) cluster of 12 virus particles seen from the three-fold axis; (b) similar cluster seen from the five-fold axis; (c) dimer cluster of 23 virus particles; (d) linear trimer cluster; (e) angled trimer cluster; (f) linear tetramer cluster.
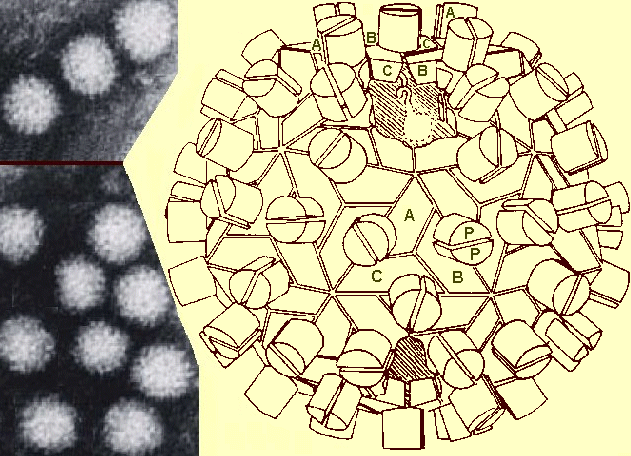
(a) Electron micrograph of negatively stained CRSV virions, (b) A presumed structural model of the virion based on the structure at 3 Å resolution of tomato bushy stunt and turnip crinkle viruses, which share significant amino acid and structural homology with all members of the Dianthovirus genus, including CRSV (Giesman-Cookmeyer et al., 1995). The model is a T=3 structure with 5:3:2 symmetry. Depending on their position in the virion, identical capsid protein subunits adopt slightly different conformational states A, B, and C (termed quasi-equivalence) to allow formation of the virion. The protruding domains (P) of the subunits are thought to account for the granular appearance of the virions in the electron microscope.

(Left) Photograph of agarose gel electrophoretic patterns of genomic ssRNA species isolated from virus particles, (right) polyacrylamide gel electrophoretic patterns of dsRNA species isolated from virus infected tissue. (A) CRSV, (B) RCNMV.
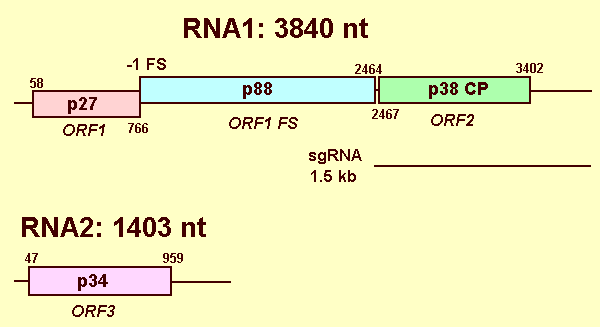
Genome organization and replication strategy. Lines represent non-coding regions and boxes represent ORFs with the sizes of the respective proteins (or readthrough products) indicated within. The nt numbers for start and stop codons defining the ORFs are indicated above and below, respectively. The -1 ribosomal frameshifting signal is identified by -1 FS. The line under RNA-1 depicts the subgenomic RNA for the 1.5 kb capsid protein.

Ultrathin section of cytoplasmic crystal composed of virus particles from inoculated cowpea leaf. Bar represents 0.5 µm.
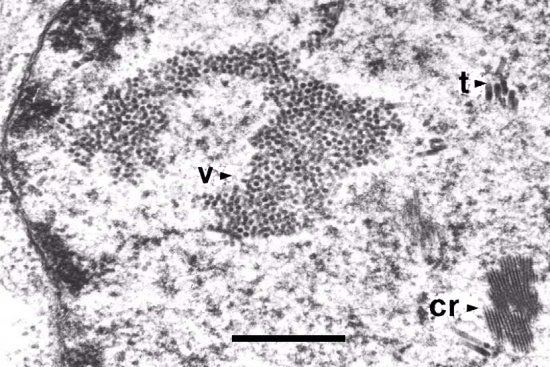
Section of infected Dianthus barbatus leaf showing nucleus containing: v, virus particles; t, tubules; and cr, protein crystal. Bar represents 0.5 µm.
References list for DPV: Carnation ringspot virus (371)
- Brown & Trudgill, Nematologica 30: 102, 1984.
- Ebbels, Plant Health, Oxford: Blackwell Scientific, 1979.
- Fritzsche & Schmelzer, Naturwissenschaften 54: 498, 1967.
- Fritzsche, Kegler, Thiele & Gruber, Archiv für Phytopathologie und Pflanzenschutz 15: 177, 1979.
- Giesman-Cookmeyer, Kim & Lommel, Dianthoviruses, in Pathogenesis and Host-Parasite Specificity in Plant Diseases: Histopathological, Biochemical, Genetic and Molecular Basis. Vol.3. Oxford: Pergamon Press. p.157, 1995.
- Hamilton & Tremaine, in The Plant Viruses, Vol. 5: Polyhedral Virions and Bipartite RNA Genomes, p. 251, ed. B. D. Harrison & A. F. Murant, New York: Plenum Press, 1996.
- Hollings & Stone, Annals of Applied Biology 56: 73, 1965.
- Hollings & Stone, CMI/AAB Descriptions of Plant Viruses 21, 3 pp., 1970.
- Hiruki, Advances in Virus Research 33: 257, 1987.
- Kalmakoff & Tremaine, Virology 33: 10, 1967.
- Kassanis, Annals of Applied Biology 43: 103, 1955.
- Kegler & Kegler, Archiv für Phytopathologie und Pflanzenschutz 17: 307, 1981.
- Kegler, Kegler & Kleinhempel, Zeszyty Problemowe Postepow Nauk Rolniczych 291: 155, 1983.
- Kegler, Verderevskaja, Proll, Fritzsche, Schmidt, Kalasjan, Kosakovskaja, Kleinhempel & Herrmann, Archiv für Phytopathologie und Pflanzenschutz 13: 297, 1977.
- Kemp, Journal of Botany 42: 45, 1964.
- Kendall & Lommel, Journal of General Virology 73: 2479, 1992.
- Kim & Lommel, Virology 200: 574, 1994.
- Kim & Lommel, Virology 250: 50, 1998.
- Kleinhempel, Gruber & Kegler, Acta Phytopathologica Academiae Scientiarum Hungaricae 15: 107, 1980.
- Koenig, An, Lesemann & Burgermeister, Journal of Phytopathology 121: 346, 1988.
- Koenig, Rudel & Lesemann, Journal of Phytopathology 2: 169, 1989.
- Koev, Kleshnina, Prizhilevskaya & Ermicheva, Sadovodstvo, Vinogradarstvo i Vinodelie Moldavii 6: 44, 1983.
- Kowalska, Phytopathologische Zeitschrift 74: 329, 1972.
- Kuhne, Leiser & Ostermann, Acta Phytopathologica Academiae Scientiarum Hungaricae 20: 109, 1985.
- Lommel, in Virus Taxonomy - 6th Report of the International Committee on Taxonomy of Viruses (Archives of Virology Suppl. 10), p. 404, eds F. A. Murphy et al., Vienna: Springer-Verlag, 1995.
- Lommel, McCain & Morris, Phytopathology 72: 1018, 1982.
- Lommel, McCain, Mayhew & Morris, Plant Disease 67: 53, 1983a.
- Lommel, Stenger & Morris, Acta Horticulturae 141: 79, 1983b.
- Noordam, Thung & Van der Want, Tijdschrift over Plantenziekten 57: 1, 1951.
- Lovisolo & Lisa, Fitopatologia Brasileira 3: 219, 1978.
- Paje-Manalo & Lommel, Phytopathology 79: 457, 1989.
- Richter, Kleinhempel, Gruber & Kegler, Archiv für Phytopathologie und Pflanzenschutz 14: 411, 1978.
- Ronald & Tremaine, Phytopathology 66: 1302, 1976.
- Rudel, Querfurth & Paul, Nachrichtenblatt des deutschen Pflanzenschutzdienstes, Stuttgart 29: 59, 1977.
- Russo, Di Franco & Martelli, Intervirology 28: 134, 1987.
- Ryabov, Generozov, Kendall, Lommel & Zavriev, Journal of General Virology 75: 243, 1994.
- Rybalko & Kharuta, Fiziologiya i Biokhimiya Kul'turnykh Rastenii 10: 535, 1978.
- Sit, Vaewhongs & Lommel, Science, N.Y. 281: 829, 1998.
- Tremaine & Dodds, CMI/AAB Descriptions of Plant Viruses 308, 5 pp., 1985.
- Tremaine & Ronald, Journal of General Virology 30: 299, 1976.
- Tremaine, Ronald & McGauley, Phytopathology 73: 1241, 1983.
- Tremaine, Ronald & McGauley, Phytopathology 74: 161, 1984.
- Tremaine, Ronald & Valcic, Phytopathology 66: 34, 1976.
- Van Ruiten, Acta Horticulturae 216: 239, 1987.
- Weintraub, Ragetli & Leung, Phytomorphology 25: 288, 1975.
- Zavriev, Hickey & Lommel, Virology 216: 407, 1996.