Details of DPV and References
DPV NO: 374 June 2000
Family: Potyviridae
Genus: Bymovirus
Species: Barley yellow mosaic virus | Acronym: BaYMV
This is a revised version of DPV 143
Barley yellow mosaic virus
M. J. Adams Plant Pathology Department, IACR-Rothamsted, Harpenden, Herts., AL5 2JQ, UK
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Ikata & Kawai (1940).
A virus with slightly flexuous filamentous particles with a diameter of c. 13 nm and two modal lengths, c. 275 and 550 nm, containing two species of ssRNA. It is transmitted by the plasmodiophorid fungus Polymyxa graminis and is sap-transmissible with difficulty. The only host known is barley. Pinwheel inclusions are formed in the cytoplasm of infected cells.
Main Diseases
Barley yellow mosaic virus is one of two agents (the other is barley mild mosaic virus) that causes yellow mosaic disease of barley. The disease is soil-borne because the viruses are transmitted by the plasmodiophorid fungus Polymyxa graminis which has resting spores that survive in soil. The viruses occur, either separately or together, in autumn-sown barley and cause similar symptoms. In the field, yellow patches appear in winter or early spring (Fig. 1). Leaves of infected plants have elongated, pale green or yellow flecks, typically on the youngest (unfurling) leaves (Fig. 2) and infected leaves may be curled, giving the plants a spiky appearance. Sometimes the leaves show complete yellowing with necrotic patches and the plants are stunted. Affected plants have fewer tillers, yield less grain and grain size may be irregular. The severity of symptoms depends on the cultivar of barley and the environmental conditions but symptoms become less obvious as the stem elongates towards flowering and upper leaves are often free of symptoms. Patches may remain visible as areas of stunted growth even when symptoms have apparently disappeared but plants sometimes appear to recover from infection. Yield losses are difficult to estimate but values of 40-80% loss in 2-rowed barley in Japan and up to 50% in Europe have been recorded (Usugi, 1988; Proeseler et al., 1988). They depend upon the barley cultivar and winter weather conditions, being greatest on crops sown early in the autumn and when the autumn is mild but the winter is especially cold (Kusaba et al., 1971).
Geographical Distribution
Widespread in Japan (Kashiwazaki et al., 1989), Eastern China (Chen et al., 1996) and various western European countries, including Belgium (Froidmont et al., 1993), England (Hill & Walpole, 1989; Adams, 1991), France (Hariri et al., 1990; Signoret & Huth, 1993), Germany (Huth, 1991; Proeseler et al., 1991) and Italy (Rubies-Autonell, 1991). It has also been reported from Greece (Katis et al., 1997), and the Ukraine (Fantakhun et al., 1987; Omel'chenko et al., 1996).
Host Range and Symptomatology
The only host known is Hordeum vulgare (barley). Transmitted through infested soil, and (with difficulty) by inoculation of sap. The species of plants tested but not infected include Triticum aestivum (wheat), Avena sativa (oat), Oryza sativa (rice), Chenopodium amaranticolor and Nicotiana tabacum (tobacco). Hordeum spontaneum and H. agriocrithon were susceptible when planted in infested soil (Miyamoto, 1958). Reports of successful mechanical inoculation of the virus to other species of Hordeum and to a few other species of the Poaceae (Proeseler, 1988) refer to what was then described as a strain of barley yellow mosaic virus but which is now recognised as barley mild mosaic virus.
Diagnostic species
Propagation species
Assay species
Strains
Reports from Europe during the period 1980-1990 refer to a strain that is readily transmissible by mechanical inoculation (BaYMV-M or BaYMV-Streatley). These isolates are now recognised as a different virus, barley mild mosaic virus (Huth & Adams, 1990).
Six different strains (in three groups) have been identified in Japan based on the response of differential cultivars: I-1, I-2, I-3, II-1, II-2 and III (Kashiwazaki et al., 1989). In Europe, two strains are recognised on the basis of the response of cultivars carrying the ym4 resistance gene. These cultivars are immune to the common strain but susceptible to a resistance-breaking strain that is usually named BaYMV-2 and which is becoming increasingly important (Huth, 1991; Hariri et al., 1990; Adams, 1991). Several strains probably occur in China but these are less well defined (Chen et al., 1996). No differences in serological or other properties have been demonstrated between any of these strains and they are distinguished only by cultivar response.
Transmission by Vectors
Transmitted by the plasmodiophorid fungus Polymyxa graminis (Kusaba et al., 1971; Adams, 1990a), which is an obligate root parasite. The virus is acquired when the plasmodia of the fungus (Fig. 3) are growing inside the barley root cells and it is transmitted within the zoospores (Fig. 4) or resting spores (Fig. 5) that it produces. The virus survives within the resting spores and infectivity has been retained in air-dried soil for 5-10 years (Usugi, 1988).
Transmission through Seed
Not found (Yasu & Yoshino, 1964).
Serology
The virus is only moderately immunogenic. Serum from a rabbit injected intravenously, then intramuscularly, with partially purified virus reacted with the virus to a titre of 1/1280 in complement fixation tests (Usugi & Saito, 1970). ISEM and ELISA are useful for routine diagnosis (Adams, 1991). Monoclonal antibodies have been prepared (Hariri et al., 1996a; 1996b).
Relationships
The different strains of BaYMV have been characterised only by cultivar response and cannot be distinguished by serological tests. Nucleotide sequencing suggests that the two European strains are extremely similar (Shi et al., 1995; 1996) and it has not so far proved possible to develop a test to distinguish them.
BaYMV resembles the other members of the genus Bymovirus in transmission by Polymyxa graminis, in particle morphology and in inducing pinwheel-type inclusions and characteristic membranous network structures in the cytoplasm of infected plant cells. There is a close serological relationship to both wheat yellow mosaic and wheat spindle streak mosaic viruses (Usugi et al., 1989) but no detectable relationship to the other viruses, including barley mild mosaic virus. The percentages of amino acids in the coat protein identical to those of the other bymoviruses that have been sequenced are: 74-76% (WSSMV), 68% (WYMV), 33-34% (BaMMV) and 34% (RNMV) (Badge et al., 1997; Namba et al., 1998).
Stability in Sap
The dilution end-point is less than 10-2 (Kusaba et al., 1971; Yasu & Yoshino, 1964). The thermal inactivation point and longevity in vitro are not known. Infectivity of dried leaf tissue is retained for two years at 5-10°C (Miyamoto, 1958).
Purification
There are several published protocols developed from the procedure of Usugi & Saito (1970). The protocol currently in use at IACR-Rothamsted is:
Homogenise 100 g leaf pieces (fresh or powdered in liquid N2 and stored at -70°C) in 400 ml cold buffer A (0.1M Tri-potassium citrate; 0.01M EDTA di-sodium salt; pH adjusted to 7.0 with KOH if necessary) + 25 ml/l Triton X-100 + 1mM phenylmethylsulfonylfluoride (PMSF; added from a 50mM stock solution in isopropanol immediately before use) and strain through muslin. Centrifuge at 15-20,000 g for 15 min at 4°C and retain supernatant. Centrifuge through a 30% (w/w) sucrose cushion at 98,000 g for 2 hours at 4°C. Resuspend pellet in cold buffer A + 1mM PMSF. Allow to resuspend for about 1 hour at 4°C then centrifuge at c. 2,500 g for 10 min at 4°C and retain supernatant. Dilute with buffer A + 1mM PMSF and repeat centrifugation through sucrose cushion, resuspend and low speed spin as before. Mix with CsCl (5.15 ml preparation + 2.85 ml stock 60% (w/w) CsCl solution for each 10 ml tube), centrifuge at 84,000 g overnight (16-17 hours) at 4°C and collect the light-scattering virus band using a bent needle and syringe. Dilute 10-20 fold in buffer A and collect by centrifugation at 67,500 g for 4 hours at 4°C.
Properties of Particles
Buoyant density in CsCl: 1.321 to 1.349 g/cm3 (Huth, 1988).
Particle weights 2.8 x 106 (RNA1) and 1.4 x 106 daltons (RNA2) (Huth, 1988)
A260/A280: 1.14 (not corrected for light-scattering).
Particle Structure
Particles (Fig. 6) of barley yellow mosaic virus are slightly flexuous filaments 13 nm in diameter and having two modal lengths of 275 and 550 nm; both lengths are found in leaf dips and in partially purified virus preparations (Inouye, 1964, 1968).
Particle Composition
Nucleic acid: Single-stranded linear RNA in two segments (RNA1, 7630-7645 nucleotides; RNA2, 3585 nucleotides) comprising about 5 % of particle weight.
Protein: A single coat protein species consisting of 297 amino acids. The mol. wt was estimated as 33-35 kDa by SDS-polyacrylamide gel electrophoresis, which is slightly larger than that (32.4 kDa) calculated from the sequence. The coat protein undergoes partial proteolysis during purification of virus particles (Ehlers & Paul, 1986; Kashiwazaki et al., 1989).
Genome Properties
Bipartite, positive-sense ssRNA with poly(A) at the 3' terminus and probably with a genome-linked protein (VPg) at the 5' terminus. There is no extensive nucleotide sequence homology between RNA1 and RNA2 except for the 5' non-coding regions (Kashiwazaki et al., 1991).
Complete nucleotide sequences have been determined for the genomes of Japanese (D01091, RNA1; D01092, RNA2) German (X69757, RNA1; D01099, RNA2) and Chinese (AJ132268, RNA1; AJ132269, RNA2) isolates. Nucleotide sequences have also been reported for the 3'-terminus (including the coat protein gene) of RNA1 of a UK isolate (Z24677) and for the coat protein coding region of three UK (X95695-X95697) and 10 Chinese isolates (AJ224619-AJ224628). Partial sequences for a region at the 5'-end of the P2 region of RNA2 have been reported for one German BaYMV-2 isolate (X69836), 3 UK isolates (X95698-X95700) and 10 Chinese isolates (AJ224629-AJ224638).
The organisation of the genome is shown in Fig. 7. RNA1 consists of 7632-7643 nucleotides, excluding the 3' poly(A), and encodes a single large polyprotein of 2410-2412 amino acids (271 kDa). Its organisation resembles the 3'-terminal three quarters of the monopartite genome of potyviruses (Kashiwazaki et al., 1990; Peerenboom et al., 1992) with the coat protein at the C-terminus and at least five, and probably seven, putative non-structural proteins which have been identified by sequence alignments and motif comparisons (Table 1). The NIa-Pro probably acts as a proteinase to catalyse cleavage of the polyprotein at the QX or EX dipeptides indicated in Fig. 7, to give rise to the functional proteins.
Table 1. Protein products of RNA1 of barley yellow mosaic virus
| kDa | Product | Function | |
| P3 | 38 | Unknown | |
| 7K | 7 | Unknown: analogous to Potyvirus 6K1 | |
| CI | 74 | Cylindrical inclusion protein | NTP-binding motif (helicase) |
| 14K | 14 | Unknown: analogous to Potyvirus 6K2 | |
| NIa-VPg | 22 | Nuclear inclusion protein | Genome-linked protein |
| NIa-Pro | 25 | Nuclear inclusion protein | Serine-like protease |
| NIb | 60 | Nuclear inclusion protein | RNA-dependent RNA polymerase |
| CP | 32 | Coat protein | Capsid protein |
RNA2 consists of 3585 nucleotides, excluding the 3' poly(A), and encodes a single polyprotein of 890 amino acids (98.2 kDa) which contains two putative functional proteins (Davidson et al., 1991; Kashiwazaki et al., 1991). P1 (28 kDa) contains a protease domain that corresponds to the helper component proteinase of potyviruses, and probably cleaves the 98K polyprotein at a GS dipeptide. By analogy with barley mild mosaic virus, P2 (70 kDa) is probably involved in fungus transmission. Both proteins have been shown to be associated with inclusion bodies in infected leaf cells (Schenk et al., 1993).
Japanese and German isolates display 94-95% nucleotide homology. Examination of European resistance-breaking BaYMV-2 isolates has shown that these are extremely similar to the common isolate: the crucial difference has yet to be identified but does not occur in the coat protein coding region (Shi et al., 1995; 1996).
Relations with Cells and Tissues
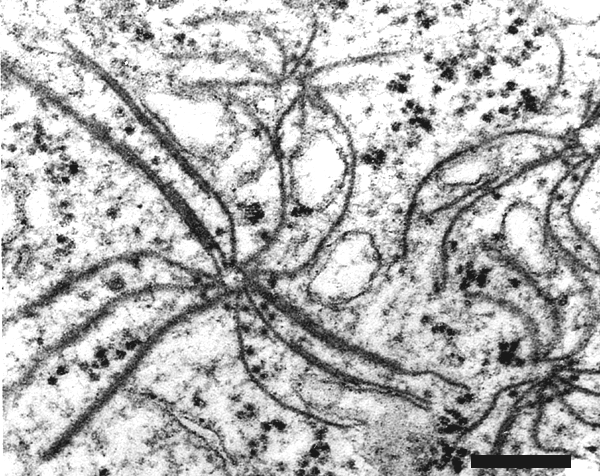
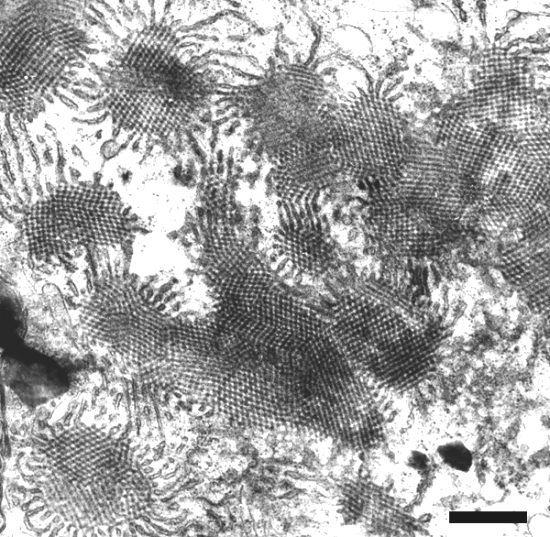
X-bodies can be found in cells of epidermal strips of infected leaves (Fig. 8). In ultrathin sections, inclusion bodies of pinwheel-type (Fig. 9), densely packed membranes (Fig. 10), and crystal-like bodies (Fig. 11) can be observed in the cytoplasm (Saito et al., 1966; Hibino et al., 1981; Schenk et al., 1993).
Ecology and Control
Because of its transmission by Polymyxa graminis, barley yellow mosaic virus is soil-borne. It survives within the fungus resting spores, which remain within root debris after the crop is harvested and can persist in soil for many years. No weed hosts of the virus are known and although some isolates of the fungus may be able to infect other cereals or grass weeds, it is generally assumed that other hosts play little or no role in the epidemiology of the disease (Adams, 1990b). There is a potential role for spring barley as a symptomless host, but this has never been demonstrated in the field.
Barley roots become infected in the autumn soon after sowing. The optimum temperature is 13-16ºC and little infection occurs below about 10ºC (Kusaba et al., 1971). The virus remains in the roots until the spring when it seems that re-growth of plants, damaged by earlier cold weather, causes the plants to mobilise root reserves, thus reversing the normal downward phloem movement. This translocates virus particles from the roots to the growing leaves where further virus multiplication results in the typical systemic mosaic symptoms (Schenk et al., 1995). The lack of symptoms on leaves produced later in the season may be related to the speed of their growth or to the higher temperatures that are unfavourable to the virus (Inouye et al., 1990). The absence of symptoms on spring-sown barley is probably due to the speed of crop development and the lack of stimulus to reverse the phloem movement.
Although general soil sterilisation may control the disease (Saito et al., 1964; Kusaba et al., 1971) it is practically difficult and economically unjustified. Other chemical treatments have not given consistent benefits (Proeseler and Kastirr, 1988). The incidence and severity of the disease can be decreased by delaying sowing in the autumn (Watanabe et al., 1989) probably because this results in less root infection by the fungus vector before the onset of winter. The value of delayed sowing as a practical control measure varies between seasons (Adams et al., unpublished) probably depending upon the autumn weather conditions. Although the disease is more severe where barley is grown continuously than where it is grown in a rotation system (one year in three or four) (Faccini et al., 1995; Rubies-Autonell et al., 1995), crop rotation is not widely practised as a control measure.
The principal means of control is the use of resistant varieties, which are available in Europe and East Asia. These varieties are susceptible to the fungus vector but are immune to the virus. Several resistance genes have been identified but some of these can be overcome by certain strains (pathotypes) of the virus or by barley mild mosaic virus. In Europe, the only widely used gene has been ym4, derived from the Yugoslav landrace Ragusa. It confers resistance to the common European strain but not to BaYMV-2 (Graner & Bauer, 1993). It also confers resistance to some, but not all, Chinese strains (Chen et al., 1996). In Japan, a wider range of resistance genes have been deployed (Kashiwazaki et al., 1989; Iida et al., 1992; Konishi et al., 1997) and some of these are now being used by European breeders.
Notes
The mosaic viruses are readily distinguished from most other viruses that affect barley by the mosaic flecking on the youngest leaves. Of those with elongated particles, soil-borne wheat mosaic virus and barley stripe mosaic virus do not usually cause the flecking symptoms, have straight rod-shaped particles and can be distinguished by serological tests. They also cause local lesions on inoculated leaves of Chenopodium amaranticolor or C. quinoa, which BaYMV does not. Barley mild mosaic virus causes identical symptoms to barley yellow mosaic virus and dual infections can also occur, especially in Europe. The two viruses can only be distinguished by serological tests (e.g. ELISA). Related, but distinct, mosaic viruses occur on other cereal crops, causing similar symptoms on oats (oat mosaic virus) and wheat (wheat yellow mosaic and wheat spindle streak mosaic viruses) in particular. Although the wheat bymoviruses have a close serological relationship to BaYMV, the viruses are confined to their respective cereal hosts.
Publications apparently describing work with BaYMV from Europe in the period 1980-1990 often used virus isolates that are now recognised as barley mild mosaic virus and this can cause confusion. In particular, the strain described from Germany as transmissible by mechanical inoculation (BaYMV-M) and from the UK as the Streatley strain are now recognised as barley mild mosaic virus (Huth & Adams, 1990).
Figures

Yellow patches in winter barley cv. Panda caused by barley yellow mosaic virus.

Mosaic leaf symptoms on barley (cv. Igri) caused by barley yellow mosaic virus.

Light micrograph of barley root stained with methylene blue, showing plasmodium of Polymyxa graminis.

Zoospore suspension of Polymyxa graminis. Main picture shows a light micrograph; inset shows a scanning electron micrograph of a single zoospore. Zoospores are about 4µm in diameter. F = flagellum.

Resting spore clusters of Polymyxa graminis: Main picture shows a light micrograph of a heavily infected barley root (not stained); inset shows a scanning electron micrograph of a single cluster. Individual spores are about 6µm in diameter.
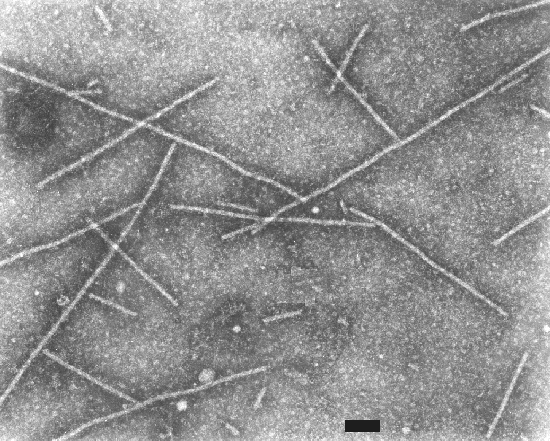
Electron micrograph of particles of barley yellow mosaic virus. Bar represents 100 nm.
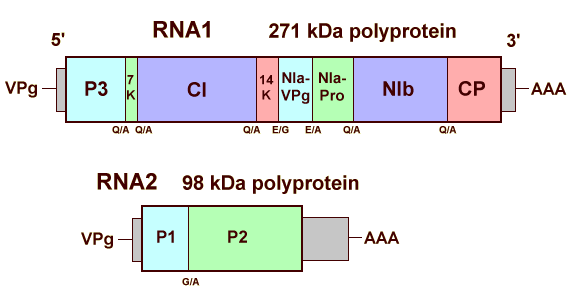
Diagram showing the genome organisation of barley yellow mosaic virus.

X-bodies in epidermal cells of barley. N = nucleus; X = X-body.
Electron micrograph of an ultrathin section of infected barley leaf showing pinwheel-type inclusions. Bar represents 100 nm. Photo courtesy of Prof. H.-H. Steinbiss, Max Planck Institut für Zuchtungsforschung, Köln, Germany.
Electron micrograph of an ultrathin section of infected barley leaf showing densely packed membranous inclusions. Bar represents 500 nm. Photo courtesy of Prof. H.-H. Steinbiss, Max Planck Institut für Zuchtungsforschung, Köln, Germany.

Electron micrograph of an ultrathin section of infected barley leaf, cv. Tapir, showing crystal-like inclusion. Bar represents 500 nm. Photo courtesy of Prof. H.-H. Steinbiss, Max Planck Institut für Zuchtungsforschung, Köln, Germany.
References list for DPV: Barley yellow mosaic virus (374)
- Adams, in: Proceedings of the First Symposium of the International Working Group on Plant Viruses with Fungal Vectors, p.121, eds R. Koening & M. J. C. Asher, Stuttgart: Ulmer, 1990a.
- Adams, Soil Use and Management 6: 184, 1990b.
- Adams, Plant Pathology 40: 53, 1991.
- Badge, Kashiwazaki, Lock & Foster, European Journal of Plant Pathology 103: 721, 1997.
- Chen, Adams, Zhu, Wang, Chen, Huang & Zhang, Plant Pathology 45: 1117, 1996.
- Davidson, Pröls, Schell & Steinbiss, Journal of General Virology 72: 989, 1991.
- Ehlers & Paul, Journal of Phytopathology 115: 294, 1986.
- Faccini, Alberici, Baravelli, Cattivelli & Delogu, Informatore Agrario 51: 59, 1995.
- Fantakhun, Pavlenko & Bobyr, Mikrobiologicheskii Zhurnal - Kiev 49: 76, 1987.
- Foroughi-Wehr, Nachrichtenblatt des Deutschen Pflanzenschutzdienstes (Stuttgart) 50: 161, 1998.
- Froidmont, Maroquin & Steyer, Parasitica 49: 107, 1993.
- Graner & Bauer, Theoretical and Applied Genetics 86: 689, 1993.
- Hariri, Fouchard & Lapierre, in Proceedings of the First Symposium of the International Working Group on Plant Viruses with Fungal Vectors, p.109, eds R. Koening & M. J. C. Asher, Stuttgart: Ulmer, 1990.
- Hariri, Delaunay, Gomes, Filleur, Plovie & Lapierre, European Journal of Plant Pathology 102: 283, 1996a.
- Hariri, Lapierre, Filleur, Ploive & Delaunay, Journal of Phytopathology 144: 331, 1996b.
- Hibino, Usugi & Saito, Annals of the Phytopathological Society of Japan 47: 510, 1981.
- Hill & Walpole, EPPO Bulletin 19: 555, 1989.
- Huth, in Developments in Applied Biology II. Viruses with fungal vectors, p.61, eds J. I. Cooper & M. J. C. Asher, Wellesbourne: Association of Applied Biologists, 1988.
- Huth, Nachrichtenblatt des Deutschen Pflanzenschutzdienstes (Stuttgart) 43: 233, 1991.
- Huth & Adams, Intervirology 31: 38, 1990.
- Iida, Watanabe, Toshima & Ogawa, Japanese Journal of Breeding 42: 863, 1992.
- Ikata & Kawai, Noji Kairyo Shiryo 154, 123 pp., 1940.
- Inouye, Nogaku Kenkyu 50: 117, 1964.
- Inouye, Annals of the Phytopathological Society of Japan 34: 301, 1968.
- Inouye, Maeda & Mitsuhata, Nogaku Kenkyu 62: 139, 1990
- Kashiwazaki, Ogawa, Usugi, Omura & Tsuchizaki, Annals of the Phytopathological Society of Japan 55: 16, 1989.
- Kashiwazaki, Minobe, Omura & Hibino, Journal of General Virology 71: 2781, 1990.
- Kashiwazaki, Minobe & Hibino, Journal of General Virology 72: 995, 1991.
- Katis, Tzavella-Klonari & Adams, European Journal of Plant Pathology 103: 281, 1997.
- Konishi, Ban, Iida & Yoshimi, Theoretical and Applied Genetics 94: 871, 1997.
- Kusaba, Toyama, Yumoto & Tatabe, Special Bulletin of the Tottori Agricultural Experimental Station 2, 208pp., 1971.
- Miyamoto, Annals of the Phytopathological Society of Japan 23: 199, 1958.
- Namba, Kashiwazaki, Lu, Tamura & Tsuchizaki, Archives of Virology 143: 631, 1998.
- Omel'chenko, Simonenko & Bondarenko, Mikrobiologichnii Zhurnal 58: 58, 1996.
- Peerenboom, Pröls, Schell, Steinbiss & Davidson, Journal of General Virology 73: 1303, 1992.
- Proeseler, Archiv für Phytopathologie und Pflanzenschutz 24: 267, 1988.
- Proeseler & Kastirr, Nachrichtenblatt für den Pflanzenschutz in der DDR 42: 116, 1988.
- Proeseler, Kegler & Henkner, Nachrichtenblatt für den Pflanzenschutz in der DDR 42: 213, 1988.
- Proeseler, Stanarius, Szigat & Herold, Nachrichtenblatt für den Pflanzenschutz in der DDR 43: 65, 1991.
- Rubies-Autonell, Difesa delle Piante 14: 43, 1991.
- Rubies-Autonell, Toderi, Marenghi & Vallega, Agronomie 15: 511, 1995.
- Saito, Takanashi, Iwata & Okamoto, Bulletin of the National Institute for Agricultural Sciences, Nishigahara, Tokyo, Series C 17: 20, 1964.
- Saito, Ueda & Inaba, Annals of the Phytopathological Society of Japan 32: 87, 1966.
- Schenk, Steinbiss, Müller & Schmitz, Protoplasma 173: 113, 1993.
- Schenk, Antoniw, Batista, Jacobi, Adams & Steinbiss, Annals of Applied Biology 126: 291, 1995.
- Shi, Zhu, Chen, Stratford, Wilson, Antoniw, Foulds, Macfarlane & Adams, Virus Research 38: 193, 1995.
- Shi, Chen, Wilson, Macfarlane, Antoniw & Adams, Virus Research 44: 1, 1996.
- Signoret & Huth, Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 100: 239, 1993.
- Takahashi, Inouye, Hayashi, Moriya, Hirao & Mitsuhata, Nogaku Kenkyu 52: 65, 1968.
- Takanashi, Saito & Iwata, Annals of the Phytopathological Society of Japan 33: 43, 1967.
- Usugi, in Developments in Applied Biology II. Viruses with fungal vectors, p.213, eds., J. I. Cooper & M. J. C. Asher, Wellesbourne: Association of Applied Biologists, 1988.
- Usugi & Saito, Annals of the Phytopathological Society of Japan 36: 375, 1970.
- Usugi T, Kashiwazaki S, Omura T, Tsuchizaki T, 1989. Annals of the Phytopathological Society of Japan 55: 26-31.
- Watanabe, Toshima, Ueda & Ogawa, Proceedings of the Kanto-Tosan Plant Protection Society 36: 30, 1989.
- Yasu & Yoshino, Bulletin of the Saitama Agricultural Experimental Station 24: 1, 1964.