Details of DPV and References
DPV NO: 377 July 2000
Family: Closteroviridae
Genus: Closterovirus
Species: Beet yellows virus | Acronym: BYV
This is a revised version of DPV 13
Beet yellows virus
Alexey A. Agranovsky Department of Virology and Belozersky Institute, Moscow State University, 119899 Moscow, Russia.
Dietrich E. Lesemann Institute of Biochemistry and Plant Virology, Federal Biological Center of Agriculture and Forestry, Messeweg 11/12, D-38104 Braunschweig, Germany.
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Roland (1936), Watson (1942) and Hull (1950).
A virus with flexuous filamentous particles c. 1350 nm long, present in beet growing areas world-wide. Natural host range is moderate, mainly in Chenopodiaceae. Transmitted by several species of aphids in a semi-persistent manner and with difficulty by mechanical inoculation of sap extracts (Bar-Joseph et al., 1979; Murant et al., 1988). Virus particles contain a single positive sense ssRNA molecule of 14.5 kb and two coat proteins of Mr 22,300 and 24,000 (p22 and p24; major and minor CP, respectively) (Agranovsky et al., 1991b; 1994a; 1994b; 1995).
Main Diseases
Causes a yellowing disease in Beta vulgaris (sugar beet, red beet, etc.) and in Spinacia oleracea (spinach).
Geographical Distribution
World-wide in sugar beet growing areas.
Host Range and Symptomatology
Infects 121 species in 15 families, although most infectible species are in the families Chenopodiaceae, Amaranthaceae, Aizoaceae, and Caryophyllaceae (Duffus, 1973).
- Diagnostic species
B. vulgaris (sugar beet). Young leaves show vein clearing and yellowing, and vein etching symptoms (Fig. 1). Older leaves show more intensive yellowing and necrotic spots (Fig. 2).
Tetragonia expansa. Clearing and yellowing of veins of younger leaves (Fig. 3), leaf yellowing and distortion of older leaves. Plants become dwarfed.
Montia (syn. Claytonia) perfoliata. In aphid-inoculated plants, systemic red-rimmed necrotic spots and chlorosis in older leaves. Local necrotic lesions in mechanically inoculated leaves but no systemic infection (Russell, 1970).
Chenopodium amaranticolor or C. quinoa. 6-7 days after mechanical inoculation with sap or purified RNA preparations, leaves develop chlorotic local lesions ca. 1 mm in diameter. Manually inoculated plants do not develop systemic infection.
Chenopodium foliosum or C. capitatum. Acute stunting, distortion and vein clearing in young leaves, and usually premature death (Russell, 1970).
Nicotiana clevelandii. Symptomless systemic infection.
- Propagation species
Tetragonia expansa is suitable for maintaining cultures and for propagating virus for virus particle purification (Kassanis et al., 1977; Rogov et al., 1993).
- Assay species
C. amaranticolor, C. quinoa and Montia (Claytonia) perfoliata are suitable local lesion hosts for mechanical inoculation assays. C. foliosum, T. expansa and B. vulgaris are suitable assay hosts for aphid inoculation.
Strains
Several strains have been described in sugar beet that cause symptoms of different severity, from mild yellowing to vein-etching and leaf necrosis (Bennet, 1960; Russell, 1970; Polack, 1971). The strains can be differentiated on C. perfoliata, C. capitatum, T. expansa and N. clevelandii; they are closely related serologically and most of them confer complete cross-protection against each other (Russell, 1970; Duffus, 1973), albeit some strains do not cross-protect (Bennet, 1960).
Transmission by Vectors
Transmitted by 22 aphid species but Myzus persicae and Aphis fabae are the principal natural vectors (Kennedy et al., 1962; Russell, 1970). Transmission is in a semi-persistent mode (Sylvester, 1956), with acquisition access and test feeding times of 12 and 6 hrs, respectively. The virus is retained by feeding aphids for 3 days, with a half-life of 8 hr (Russell, 1970). Not transmitted to progeny vectors and not retained after moulting (Watson, 1960).
Transmission through Seed
Not transmissible through seed and pollen, despite some earlier reports.
Transmission by Dodder
Several species can transmit (Fuchs & Beiss, 1954). Aphids can acquire virus from some non-transmitting species of dodder growing on infected plants (Bennet, 1960).
Serology
Moderately immunogenic in rabbits (Bercks & Zimmer, 1956; Fuchs et al., 1979; Roseboom & Peters, 1984). Polyclonal antisera were produced with titres up to 1.9 x106 in indirect ELISA, suitable for detecting virus in beet leaves, roots, and in viruliferous aphids (Rogov et al., 1993). Rabbit polyclonal and mouse monoclonal antibodies were produced against bacterially-expressed CPs p22 and p24 (Agranovsky et al., 1994b, 1995) and against recombinant non-structural proteins encoded by the virus (Agranovsky et al., 1997; Erokhina et al., 2000).
Relationships
No serological relationships detected to other viruses (Brandes & Berks, 1965). With respect to the genome structure and amino acid sequences encoded, the virus is most closely related to members of the genus Closterovirus (family Closteroviridae), namely Citrus tristeza virus, Beet yellow stunt virus, and Grapevine leafroll-associated virus-5 (Agranovsky, 1996; Karasev et al., 1995, 1996; Zhu et al., 1998). The earlier reports on the relatedness to Apple stem grooving virus and Apple chlorotic leafspot virus were disproved by genome sequence analysis (Agranovsky et al., 1991a; 1994a).
Stability in Sap
Thermal inactivation point (10 min) in sap is 55°C. Dilution end point is up to 10-4. Infectivity is retained in frozen sap for more than a year, but is rapidly lost at 20°C (Russell, 1970).
Purification
The procedure of Kassanis et al. (1977) modified by Rogov et al. (1993) yields 15-30 mg of purified virus particles from 100 g of infected T. expansa leaves . Harvest systemically infected young leaves 3 to 7 weeks after inoculation and homogenise with a mortar and pestle in 3 volumes (w/v) of 0.1 M ammonium acetate buffer, pH 7.2 containing 0.02 M EDTA and 0.02 M DIECA. This and further procedures are done at +4°C or in wet ice. Filter through cheesecloth and centrifuge sap for 15 min at 5,000 g. Add Triton X100 to 2.5% and gently agitate the mixture for 30 min. Pellet the virus by centrifugation for 90 min at 100,000 g and resuspend the pellet in 1/20 of the initial sap volume of 0.01 M sodium borate buffer pH 7.8 (buffer B) overnight. Clarify the suspension by centrifuging for 10 min at 8,000 g and layer it over 5 ml of a sucrose cushion (30% sucrose, 7.5% PEG (mol. wt 6,000), and 0.1 M NaCl in buffer B) in a 30-ml ultracentrifuge tube. Pellet the virus by centrifugation for 120 min at 100,000 g. Briefly rinse the pellet with distilled water, resuspend in buffer B, and clarify by low speed centrifugation. Repeat the high and low speed centrifugation steps. Resuspend the final pellet in 2-3 ml of buffer B and store at +4°C, or add 50% glycerol for longer storage at -20°C. The viral RNA tends to degrade upon phenol extraction from purified virus particles suspended in borate buffer; therefore for RNA extraction, resuspend the virus particles in 0.02 M sodium phosphate, pH 7.0.
Properties of Particles
Virus particles have a sedimentation coefficient S20, w of 110 to 130 and a buoyant density in CsCl of 1.33-1.34 g.cm-3 (Bar-Joseph & Hull, 1974; Kassanis et al., 1977). Absorbance at 260 nm is 2.5 (1 mg.ml-1, 1 cm light path; corrected for light scattering);. The UV absorption spectrum of the purified particles is anomalous, with A260/A280 of 1.44 (Kassanis et al., 1977). Virus particles are rather unstable and sensitive to RNase.
Particle Structure
Virus particles are very flexuous filaments of 1250-1450 nm in length and 10 nm in diameter (Fig. 4), with a core of 3-4 nm (Leyon, 1951; Brandes & Zimmer, 1955; Mundry, 1958; Bar-Joseph et al., 1979). The helical structure of particles consists 8.5 subunits of CP per turn, and repeated every two turns (Bar-Joseph et al., 1972; Chevallier et al., 1983). The particles are built of two related CPs, p24 and p22, that form the respective virion segments of 75 nm and 1290 nm (Fig. 5 & Fig. 6) consisting of approximately 200 and 3800 subunits per RNA molecule (`rattlesnake' structure; Agranovsky et al., 1995). The tail segment has a distinct helical pitch of 4.05 nm, whereas the main part of the virion has a pitch of 3.45 nm (Fig. 5; Hills & Gay, 1976). The p24 encapsidates a 5'-terminal region of the viral RNA (Zinovkin et al., 1999). The p24-coated tail is prone to dissociate in crude sap and is easily lost during virus particle purification (Agranovsky et al., 1995; Lesemann, unpublished observation).
Particle Composition
Nucleic acid: A single species of a linear positive sense ssRNA comprising 5-6% of the particle weight (Bar-Joseph & Hull, 1974; Carpenter et al., 1977). The RNA is likely to be capped and contains no VPg, poly(A) or tRNA-like structure (Karasev et al., 1989; Agranovsky et al., 1991b, 1994a). The sequences show a slight bias in nucleotide frequencies, being uridine-rich (29%).
Protein: In PAGE of protein from purified particle preparations a single protein of Mr c.22 kDa (p22) is detected (Bar-Joseph & Hull, 1974; Carpenter et al., 1977). A 24-kDa protein (p24), present in minor amounts (c.1 molecule per 17 molecules of p22), escapes detection by protein electrophoresis (Agranovsky et al., 1995). p22 has a low content of aromatic amino acids, which accounts for the anomalous UV absorption spectrum of virus particles (Bar-Joseph & Hull, 1974; Carpenter et al., 1977; Kassanis et al., 1977). p22 is stable to degradation upon storage of purified particle preparations at 4°C (Carpenter et al., 1977).
Genome Properties
The complete nucleotide sequences are available for a Ukraine isolate (15, 480 nt; Agranovsky et al., 1991a, 1994a; accession no. X73476) and a California isolate (Peremyslov et al., 1998; accession no. AF056575). These show 99.4% overall homology (Peremyslov et al., 1998). The genome comprises a single linear molecule of positive sense ssRNA which contains nine ORFs (Fig. 7). The 5'-terminal two-thirds of the sequence contain partially overlapping ORFs 1a and 1b, encoding products with a calculated Mr of 295 and 48, respectively. The 1a product encompasses the conserved domains of a leader papain-like cysteine proteinase, methyltransferase, and helicase, whereas the 1b product contains the RNA-dependent RNA-polymerase domain (Agranovsky et al., 1991b, 1994a). ORFs 2, 3, 4, 5, 6, 7 and 8 encode proteins of, respectively, 6.4 kDa (hydrophobic protein, p6), 65 kDa (p65, homologous to the HSP70 family of cell chaperones), 64 kDa (p64), 24 kDa (p24, CP), 22 kDa (p22 CP), 20 kDa (p20) and 21 kDa (p21) (Agranovsky et al., 1991a; 1991b; 1994a; Boyko et al., 1992). The array of replication-associated domains encoded in ORFs1a and 1b is conserved in the alphavirus-like superfamily of eukaryotic viruses; the proteinase domain encoded in ORF 1a and the products of ORFs 2-6 are conserved in related closteroviruses; ORFs 7 and 8 encode unique proteins (Agranovsky, 1996; Dolja et al., 1994). The functions of some viral proteins in infection were identified. The 1a and 1b domains are necessary and sufficient for RNA replication; the leader proteinase and the p21 serve as enhancers of RNA amplification (Peremyslov et al., 1998). p65 has properties of ATPase in vitro (Nikiforova et al., 1995; Agranovsky et al., 1997) and the activity of a viral movement protein in vivo (Agranovsky et al., 1998; Peremyslov et al., 1999). p6, p64, p24 and p22 are also essential for the virus cell-to-cell movement (Alzhanova et al., 2000).
The genome is expressed by an apparently complex strategy, of which the following key elements are identified: (i) ORF 1b is likely to be expressed as a 1a/1b fusion product resulting from +1 ribosomal frameshifting (Agranovsky et al., 1994a); (ii) the proteinase domain mediates the release of a N-terminal 66-kDa leader protein from the 295-kDa 1a polyprotein, and the C-terminal part of the 1a product is processed further into at least 3 fragments (Agranovsky et al., 1994a, Agranovsky, 1998; Erokhina et al., 2000); (iii) internal genes (ORFs 2-8) are expressed via a nested set of at least six subgenomic RNAs (Dolja et al., 1990; Agranovsky et al., 1994b; Xe et al., 1996).
Relations with Cells and Tissues
The virus is confined mainly to phloem tissue, where massive particle aggregates are produced (Fig. 8), often forming banded inclusions (Esau & Hoefert, 1971b). Virus infection induces the formation of characteristic vesicles that occur in groups within membrane compartments (Fig. 9; Esau & Hoefert, 1971a; Lesemann, 1988). These structures, referred to as Beet yellows virus-type (BYV-type) vesicles, are considered of diagnostic value for BYV and the other members of the family Closteroviridae (Lesemann, 1988). Vesicles contain fine fibrils resembling double stranded nucleic acid (inset in Fig. 9; Essau & Hoefert, 1971a) and the methyltransferase-like and helicase-like proteins encoded in ORF 1a (Erokhina, Vitushkina, Lesemann, Jelkmann & Agranovsky, unpublished data), suggesting that virus replication is associated with vesicle membranes.
Ecology and Control
Beet yellows disease is responsible for severe yield losses in sugar beet, estimated at 2 to 5 per cent for every week plants are infected (Duffus, 1973; Bar-Joseph et al., 1979). The virus is not seed-borne and does not spread by contact. Because of its semi-persistent mode of transmission by aphids, the virus has a potential to spread over long distances. However, the virus spreads predominantly to areas adjacent to virus sources (Falk & Duffus, 1988). The disease incidence correlates with the presence of overwintering beets in the field and with the numbers of Myzus persicae present (Falk & Duffus, 1988). Measures to control the disease involving cultural practices, such as, the elimination of overwintering beets, early sowing, and dense spacing, as well as application of insecticides to control aphids, have been effective (Duffus, 1973; Bar-Joseph et al., 1979).
Notes
Beet yellows virus and Beet yellow stunt virus (Duffus, 1973) have several characteristics in common. Both viruses are transmitted in a semi-persistent manner by aphids, cause yellowing diseases of sugar beet and C. capitatum (C. foliosum), have flexuous filamentous particles containing genomic RNA of apparently similar size, and their infections result in similar cytopathological effects. However, they differ markedly in host range, especially in Asteraceae (Sonchus oleraceus and Lactuca sativa are immune to Beet yellows virus), in geographical distribution (Beet yellow stunt virus occurs only in California), and physicochemical and serological properties (Reed & Falk, 1989). The genomes of these two viruses are sufficiently distinct to classify them as different viruses (Karasev et al., 1996). Beet yellows virus often occurs in beet together with Beet mild yellowing luteovirus. However, these two viruses are remote with respect to the genome structure and proteins encoded, and also differ in their mode of transmission by aphids, symptomatology (Beet mild yellowing virus does not induce necrosis in chlorotic areas of the leaf), host range, particle morphology and serology (Russell, 1970).
Figures

Yellow vein etching in a young leaf of field-infected sugar beet.

Leaf yellowing and necrotic spots in an old leaf of field-infected sugarbeet.

Vein yellowing in an experimentally infected leaf of Tetragonia expansa.

General morphology of the BYV particles in crude extracts from infected Tetragonia expansa negatively stained with aqueous uranyl acetate pH 4.3; bar represents 1 µm.

Fine structure of BYV particles in crude extracts from infected Tetragonia expansa negatively stained with aqueous uranyl acetate pH 4.3, exhibiting a partially dissociated end with a larger periodicity of cross banding; bar represents 300 nm.

Serology of the BYV particles in crude extracts from infectedTetragonia expansa negatively stained with aqueous uranyl acetate pH 4.3. (a) Decoration plus immunogold labelling of a virus particle with antiserum specific to the p22 (major CP); note nonlabelled 'p24-tail'. (b) Specific decoration by antiserum specific to the p24 (minor CP) at the 75 nm 'tail' on a complete particle. (c) Selected particle ends showing specific decoration or immunogold labelling, respectively, of the 75 nm 'tails' by the antiserum specific to the p24. Bar represents 300 nm.
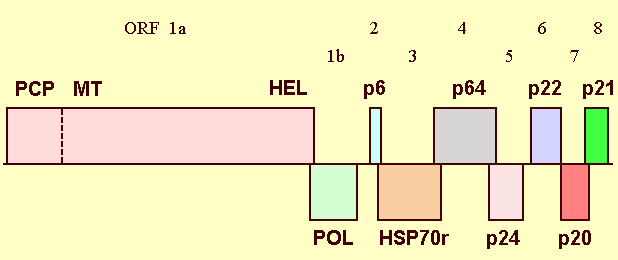
Genome structure of BYV. ORFs are shown as boxes. PCP, papain-like cysteine proteinase (with the cleavage site indicated by dotted line); MT, methyltransferase; HEL, helicase; POL, RNA-dependent RNA polymerase; HSP70r, HSP70-related 65-kDa (movement) protein; p24 and p22, minor and major capsid proteins, respectively.
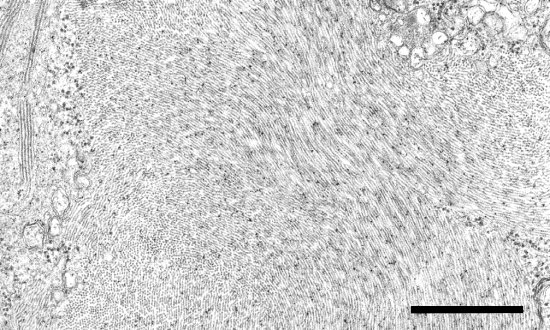
Cytological alterations induced by BYV in the infected cells of Chenopodium foliosum: massive aggregates of filamentous particles in a phloem parenchyma cell. Bar represents 1 µm.
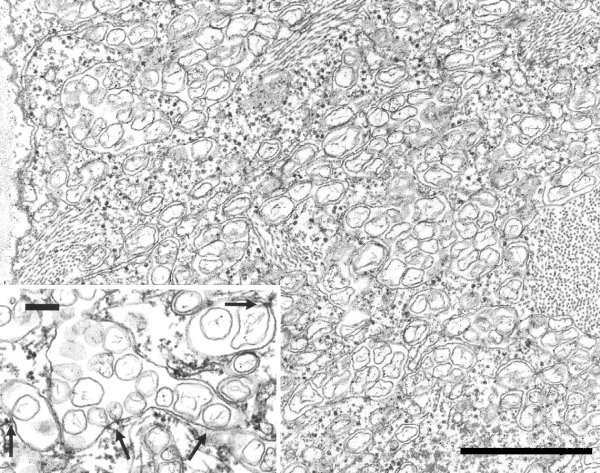
Cytological alterations induced by BYV in infected cells of Chenopodium foliosum: BYV-type vesicles in a leaf parenchyma cell (bar represents 1 µm). Inset presents details of clusters of c.100-200 nm diameter 'pseudo-spherical' vesicles containing irregular fibrous strands (bar represents 200 nm). The strands resemble structures proven to be dsRNA in similarly structured vesicles induced by other positive sense ssRNA-containing viruses (e.g. members of the genus Tombusvirus). The vesicles originate from budding into a membrane-surrounded compartment of unknown origin at the points indicated by arrows.
References list for DPV: Beet yellows virus (377)
- Agranovsky, Advances in Virus Research 47: 119, 1996.
- Agranovsky, in Handbook of Proteolytic Enzymes, p. 700, eds A.J. Barrett, N.D. Rawlings & J.F. Woessner, Academic Press, NY, 1998.
- Agranovsky, Boyko, Karasev, Koonin, & Dolja, Journal of Molecular Biology 217: 603, 1991a.
- Agranovsky, Boyko, Karasev, Lunina, Koonin, & Dolja, Journal of General Virology 72:, 15-23, 1991b.
- Agranovsky, Koonin, Boyko, Maiss, Froetschl, Lunina & Atabekov, Virology 198: 311, 1994a.
- Agranovsky, Koenig, Maiss, Boyko, Casper, & Atabekov, Journal of General Virology 75: 1431, 1994b.
- Agranovsky, Lesemann, Maiss, Hull, & Atabekov, Proceedings of the National Academy of Sciences, U.S.A. 92: 2470, 1995.
- Agranovsky, Folimonova, Folimonov, Denisenko, & Zinovkin, Journal of General Virology 78: 535, 1997.
- Agranovsky, Folimonov, Folimonova, Morozov, Schiemann, Lesemann, & Atabekov, Journal of General Virology 79: 889, 1998.
- Alzhanova, Hagiwara, Peremyslov & Dolja, Virology 268: 192, 2000.
- Bar-Joseph, Garnsey & Gonsalves, Advances in Virus Research 25: 93, 1979.
- Bar-Joseph & Hull, Virology 62: 552, 1974.
- Bar-Joseph, Loebenstein & Cohen, Virology 50: 821, 1972.
- Bennet, Technical Bulletin of the US Department of Agriculture, 1218, 1960.
- Berks & Zimmer, Journal of Phytopathology 25: 255, 1956.
- Boyko, Karasev, Agranovsky, Koonin, & Dolja, Proceedings of the National Academy of Sciences, U.S.A. 89: 9156, 1992.
- Brandes and Bercks, Advances in Virus Research 11: 1, 1965.
- Brandes & Zimmer, Journal of Phytopathology 24: 211, 1955.
- Carpenter, Kassanis & White, Virology 77: 101, 1977.
- Chevallier, Engle, Wurtz & Charles, Journal of General Virology 64: 2289, 1983.
- Dolja, Karasev & Agranovsky, New Aspects of Positive-Strand RNA Viruses, p. 31 (eds M. Brinton & F. Heinz), Washington, DC: ASM Publications, 1990.
- Dolja, Karasev & Koonin, Annual Review of Phytopathology 32: 261, 1994.
- Duffus, Advances in Virus Research 18: 347, 1973.
- Erokhina, Zinovkin, Vitushkina, Jelkmann, & Agranovsky, Journal of General Virology 81: 597, 2000.
- Esau & Hoefert, Protoplasma 72:, 255, 1971a.
- Esau & Hoefert, Protoplasma 72:, 459, 1971b.
- Falk & Duffus, in The Plant Viruses, Vol. 4: The Filamentous Plant Viruses, p. 275, (ed. R. G. Milne), NY: Plenum Press, 1988.
- Fuchs & Beiss, Naturwissenschaften 41: 506, 1954.
- Fuchs, Opel & Hartleb, Archiv fuer Phypathologie und Pflanzenschutz, Berlin 15: 73, 1979.
- Hills & Gay, John Innes Institute Annual Report, 67: 107, 1976.
- Hull, Bulletin of the Ministry of Agriculture and Fisheriesng, London 142, 52 pp., 1950.
- Karasev, Agranovsky, Rogov, Miroshnichenko, Dolja & Atabekov, Journal of General Virology 70: 241, 1989.
- Karasev, Boyko, Gowda, Nikolaeva, Hilf, Koonin, Niblett, Cline, Gumpf, Lee, Garnsey, Lewandowski & Dawson, Virology 208: 511, 1995.
- Karasev, Nikolaeva, Mushegian, Lee & Dawson, Virology 221: 199, 1996.
- Kassanis, Carpenter, White & Woods, Virology. 77: 95, 1977.
- Kennedy, Day & Eastop, A conspectus of aphids as vectors of plant viruses, London, Commonwealth Institute of Entomology, 1962.
- Lesemann, in The Plant Viruses, Vol. 4: The Filamentous Plant Viruses, p. 179, (ed. R. G. Milne), Plenum Press, NY, 1988.
- Leyon, Arkiv fuer kemi, mineralogi och geologi (Stockholm) 3: 105, 1951.
- Mundry, Zeitschrift Naturforschung 13b: 19, 1958.
- Murant, Raccah & Pirone, in The Plant Viruses, Vol. 4: The Filamentous Plant Viruses, p. 237, (ed. R. G. Milne), Plenum Press, NY, 1988.
- Nikiforova, Agranovsky & Atabekov, Doklady Rosiiskoi Akademii Nauk 340: 416, 1995.
- Peremyslov, Hagivara & Dolja, Journal of Virology 72: 5870, 1998.
- Peremyslov, Hagivara & Dolja, Proceedings of the National Academy of Sciences, U.S.A. 96: 14771, 1999.
- Polack, Journal of Phytopathology 72: 235, 1971.
- Reed & Falk, Plant Disease 73: 358, 1989.
- Rogov, Karasev & Agranovsky, Journal of Phytopathology 137: 79, 1993.
- Roland, La sucrerie belge (Bruxelles) 55: 313, 1936.
- Rozeboom & Peters, Netherland Journal of Plant Pathology 87: 133, 1984.
- Russell, CMI/AAB Descriptions of Plant Viruses 13, 1970.
- Sylvester, Journal of American Society of Sugar Beet Technology 9: 56, 1956.
- Watson, Annals of Applied Biology 29: 358, 1942.
- Watson, Reports of the 7th Commonwealth Entomology Conference, London: 157, 1960.
- Xe, Rao & Creamer, Phytopathology 87: 347, 1996.
- Zhu, Ling, Goszczynski, McFerson & Gonsalves, Journal of General Virology 79: 1289, 1998.
- Zinovkin, Jelkmann & Agranovsky, Journal of General Virology 80: 269, 1999.