Details of DPV and References
DPV NO: 378 April 2001
Family: Secoviridae
Genus: Comovirus
Species: Cowpea mosaic virus | Acronym: CPMV
This is a revised version of DPV 197
Cowpea mosaic virus
A. van Kammen Laboratory of Molecular Biology, Wageningen University, Wageningen, The Netherlands
J. van Lent Laboratory of Virology, Wageningen University, Wageningen, The Netherlands
J. Wellink Laboratory of Molecular Biology, Wageningen University, Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Chant (1959) and Van Hoof (1963).
- Synonyms
- Cowpea yellow mosaic virus (Chant, 1959)
- Cowpea mosaic virus, yellow strain (Agrawal, 1964)
An RNA-containing virus with isometric particles about 28 nm in diameter. It has a limited host range, is transmitted mainly by beetles and readily by sap inoculation. Infected plants contain two kinds of nucleoprotein particle similar in size but differing in RNA content. Particles containing no RNA are also produced by most isolates. The RNA species in different particle types represent separate parts of the viral genome.
Main Diseases
One of the most commonly reported virus diseases of cowpea (Vigna unguiculata), in which it causes mosaic and decreases leaf area and flower production. Yield reductions up to 95% have been reported. Late infections had less effect on yield than early ones (Chant, 1960). Also found in soybean (Glycine max), but causes little harm (Thottappilly & Rossel, 1985). Bock (1971) reported the susceptibility of pigeon pea (Cajanus cajan) which is widely grown in the coastal areas of Kenya as a perennial crop and may serve as a reservoir of the virus.
Geographical Distribution
Reported in Africa from field collected plants in Nigeria, Kenya, Tanzania, Togo, Mali and the Republic of Benin (Thottappilly & Rossel, 1985; Thottappilly, 1992); also from the Philippines (Talens, 1979) and Iran (Kaiser et al., 1968). Is apparently absent from North and South America (Valverde & Fulton, 1996), where in contrast cowpea severe mosaic virus (CPSMV) is common. The origin of the CPMV isolate reported from Surinam (Agrawal, 1964) needs to be confirmed.
Host Range and Symptomatology
The host range is rather limited; few hosts are known outside the Leguminosae. Individual isolates of CPMV may differ in host range and in severity of symptoms. Almost all host species show necrotic or chlorotic lesions in inoculated leaves. Cowpea varieties differ greatly in type of reaction and severity of symptoms. Immunity, tolerance and hypersensitivity occur. Symptoms in susceptible varieties may range from a hardly discernible green mottle to distinct yellow mosaic and distortion with significantly reduced growth (Bliss & Robertson, 1971).
- Diagnostic species
Vigna unguiculata (cowpea) cv. California Blackeye. Chlorotic spots with diffuse borders (diam. 1-3 mm) are produced in inoculated primary leaves (Fig. 1). Trifoliolate leaves develop a bright yellow or light green mosaic (Fig. 2) of increasing severity in younger leaves. Distortion and reduction in size are moderate. Chlorotic spots are produced on pods (Fig. 3). Plants do not show necrosis.
Chenopodium amaranticolor. In inoculated leaves, yellow local lesions (diam. 0.5-1 mm) later becoming necrotic (Fig. 4); Systemic symptoms are severe mosaic, chlorotic spots, distortion and puckering.
- Propagation species
Vigna unguiculata cv. California Blackeye is a good source of virus for purification and for maintaining cultures.
- Assay species
Phaseolus vulgaris cv. Pinto is a suitable local lesion host. Virus can easily be transferred from local lesions due to their high virus content.
Strains
Little systematic work has been done to compare isolates of the virus from different parts of the world. However, isolates from field-collected plants from Surinam (SB-isolate) and Nigeria do not seem to differ in host range and are closely related serologically (Agrawal & Maat, 1964), although there are some differences between host ranges of the Nigerian and the Kenyan isolates (Bock, 1971). In the laboratory, Bruening (1969) isolated from the SB-strain two naturally occurring variants which differed from the parent strain in the relative amount of RNA-free particles produced. De Jager (1973) isolated from the SB-strain two naturally occurring variants which, unlike the parent strain, infected Early Red cowpeas systemically.
Transmission by Vectors
Transmitted by various beetles with biting mouthparts. In Africa the chrysomelid beetle Ootheca mutabilis is an efficient vector (Chant, 1959; Bock, 1971) but Paraluperodes quaternus (Chrysomelidae) and Nematocerus acerbus (Curculionidae) were also found to transmit the virus (Whitney & Gilmer, 1974). Jansen & Staples (1971) listed Cerotoma trifurcata, Diabrotica balteata, D. undecimpunctata howardi, D. virgifera and Acalymma vittatum (all chrysomelid beetles) as vectors. The transmission is characterised by short acquisition and inoculation access periods and an apparent lack of a latent period (Gergerich and Scott, 1996). Beetle vectors may remain viruliferous for 1-2 to more than 8 days depending on the species (Chant, 1959; Jansen & Staples, 1971). Transmission efficiency and retention of infectivity are correlated with the amount of vector feeding (Jansen & Staples, 1971). Whitney & Gilmer (1974) reported also transmission by two species of thrips (Sericothrips occipitalis and Taeniothrips sjostedti) and by two species of grasshoppers (Cantotops spissus spissus and Zonocerus variegatus) but these results need to be confirmed.
Transmission through Seed
Gilmer et al., (1974) reported 1-5% seed transmission in cowpea in Nigeria, but Thottappilly & Rossel (1988) found no evidence of seed transmission using many seeds of different cowpea varieties.
Serology
The virus is strongly immunogenic. Standard methods using rabbits give antisera with high titres. Mouse monoclonal antibodies have been raised and characterized (Porta et al., 1994). Virus preparations tested by the Ouchterlony method give a single band of precipitate, even though they contain more than one electrophoretic component.
Relationships
The criteria demarcating species in the genus Comovirus are: coat protein and polymerase amino acid sequences less than 80% identical, and no pseudo-recombination between components possible (Wellink et al., 2000). A weak serological relationship is reported between cowpea mosaic virus and some other viruses of the genus Comovirus i.e. cowpea severe mosaic virus (Swaans & Van Kammen, 1973); bean pod mottle virus (Bancroft, cited by Agrawal & Maat, 1964); red clover mottle virus (Agrawal, 1964); broad bean true mosaic virus (Jones & Barker, 1976); bean rugose mosaic virus (Gamez, 1972).
Stability in Sap
Properties in vitro reported for the various isolates vary considerably, probably because of the use of different source plants and/or assay hosts. Dilution end-points range from 10-4.7 to 10-6.7. Thermal inactivation points were reported from 55 to 65°C. Longevity in vitro was found to vary from 4 to 10 days.
Purification
Yields of virus may reach 2 g/kg leaf tissue from Vigna plants grown at 30°C in a growth chamber. Freshly harvested leaves are homogenized with twice their weight of 0. 1 M phosphate buffer, pH 7.0. The homogenate is squeezed through two layers of Miracloth and centrifuged at 15,000 g for 20 min. The supernatant fluid is kept, and the pellet is washed once by suspending in phosphate buffer (0.25 ml/g leaf tissue) followed by centrifugation at 15,000 g. The combined supernatant fluids are stirred for 1 min with 0.7 volume of a 1:1 mixture of chloroform and n-butanol. After low speed centrifugation, the clear aqueous layer is removed, and the virus precipitated by adding polyethylene glycol 6000 (PEG) to 4% and NaCl to 0.2 M, and stirring for 60 min at room temperature. The precipitate after centrifugation at 15,000 g for 15 min is suspended in phosphate buffer (0.5 ml/g leaf tissue). After centrifugation at 27,000 g for 15 min, the supernatant fluid containing the virus is kept and the pellet is extracted once more in a few ml of phosphate buffer and then centrifuged at 27,000 g for 15 min. The combined supernatant fluids are layered on top of 1 ml of 40% (w/v) sucrose in 0.1 M phosphate buffer in a centrifuge tube and then centrifuged at 150,000 g for 2.5 h. The clear virus pellet is dissolved in sterile double-distilled water and centrifuged at 10,000 g for 15 min to remove possible contaminants. This procedure (Klootwijk et al., 1977) provides highly purified virus. It is a combination of the older methods of purification using chloroform/butanol mixtures (Bruening & Agrawal, 1967) or PEG and NaCl (Van Kammen, 1967).
Properties of Particles
Purified preparations of virus contain three centrifugal components: empty protein shells without RNA (T) and two nucleoprotein components (M and B), containing 24% and 34% RNA respectively. The separated nucleoprotein components are not infective (Van Kammen, 1968), but mixtures of the M and B components are. The infectivity of a mixture depends upon the proportions of the two components and the concentration of the component present in the lowest amount. Mixtures of the M or B components with the B or M components of mutants of cowpea mosaic virus are infective (Bruening, 1969; De Jager & Van Kammen, 1970; De Jager, 1976) but heterologous mixtures of components of cowpea mosaic virus and those of other comoviruses are not.
Sedimentation coefficients (s20,w) at infinite dilution (svedbergs): 58 (T), 98 (M), 118 (B). The proportion of M:B is about 1:1.
Molecular weight: 3.94 x 106 (T), 5.16 x 106 (M) and 5.98 x 106 (B).
Isoelectric point: between pH 3.7 and 4.5.
Absorbances at 260 nm (1 mg/ml, 1 cm light path): 6.2 (M), 10.0 (B), at 280 nm (1 mg/ml, 1 cm light path): 1.28 (T).
A260/A280: 1.64 (unfractionated virus), 1.61 (M), 1.74 (B).
Buoyant density in CsCl (g/cm-3) in 0.1 M phosphate buffer: 1.29 (T), 1.40 (M); component B gives two bands 1.42 (B-upper) and 1.47 (B-lower). The higher density of B-lower is the result of Cs+ ions displacing polyamines associated with the RNA (Virudachalam et al., 1985). The two bands have the same sedimentation coefficient (Bruening, 1969).
Purified preparations of the virus contain two electrophoretic components, each of which contains all three centrifugal components (Semancik, 1966). The electrophoretic forms can be separated by electrophoresis in a sucrose gradient. Electrophoretic mobility: -4.0 to -4.25 x 10-5 and -2.6 to -2.8 x 10-5 cm 2 sec-1 volt-1 in 0.1 M phosphate buffer. The slower migrating form predominates in early infection and the faster one in late infection. The slower migrating form is converted into the faster migrating form in vivo by loss of a peptide of 24 amino acids from the carboxyl-terminus of the smaller capsid protein (Taylor et al., 1999). The conversion of the slower form into the faster form can be achieved in vitro by incubation with proteolytic enzymes. The two electrophoretic forms of the virus have similar infectivity (Niblett & Semancik, 1969; Geelen et al., 1973).
Particle Structure
Particles are icosahedral with 5:3:2 axial symmetry, and a diameter of 28 nm (Fig. 5). Virion structure has been determined to 2.8 Å resolution (Lin et al., 1999). The particles consist of 60 copies each of three different wedge-shaped ß-sandwich structures arranged in a pseudo T=3 lattice. The large capsid protein (L) comprises two ß-sandwich folds and is clustered about the 3-fold axes, and the small capsid protein (S) comprises the third ß-sandwich fold and forms pentamers around the 5-fold symmetry axes.
Particle Composition
Nucleic acid: RNA, single-stranded. Components M and B each contain a single RNA molecule of 3481 (RNA2) and 5889 (RNA1) nucleotides respectively, excluding poly A tails 150-200 residues long at their 3'-ends (El Manna & Bruening, 1973). Both RNA strands contain a small protein (VPg) at their 5'-end (Daubert et al., 1978; Stanley et al., 1978).
Protein: The protein shell of the virus consists of 60 copies each of two proteins of M. Wt. 41,249 (L) and 23,708 (S) (Van Wezenbeek et al., 1983). A report that the coat proteins contain about 1.9% carbohydrates covalently linked (Partridge et al., 1974) has been disproved (Altmann & Lomonossoff, 2000).
Polyamines: Purified virus contains 5.05 µg spermidine and 0.17 µg spermine per mg virus (Nickerson & Lane, 1977). Top component contains no polyamines; M and B-upper components contain about 200 polyamine molecules per particle.
Genome Properties
The genome comprises RNA1 (accession no X00206; Lomonossoff & Shanks, 1983) and RNA2 (accession no X00729; Van Wezenbeek et al., 1983). Both genome RNAs contain a small protein (VPg) at their 5'-end and a poly A tail at their 3'-end (El Manna & Bruening, 1973; Daubert et al., 1978; Stanley et al., 1978). Each RNA species contains one large open reading frame, and they are translated in vitro as well as in vivo into one (RNA1) or two (RNA2) polyproteins that are cleaved by a viral proteinase (encoded by RNA1) to give 15 intermediate and final processing products (Fig. 6) (Goldbach and Wellink, 1996). RNA1 carries all the information for RNA replication, including the polymerase, VPg and a protein containing a nucleotide binding site, whereas RNA2 codes for the two capsid proteins and the movement protein, which are involved in cell-to-cell transport of the virus.
Relations with Cells and Tissues
Inclusion bodies can be observed in virus-infected cells by the light microscope, after staining with phloxine, as a red amorphous mass near or surrounding the nucleus (Agrawal, 1964; Swaans & Van Kammen, 1973). Electron micrographs of virus-infected cells show cytopathic structures, often adjacent to the nucleus, consisting of arrays of vesicles forming a kind of reticulum, the spaces between the vesicle areas being filled with electron-dense material that does not seem to have obvious structure (Fig. 7). The vesicles often contain fibrillar material of unknown nature (De Zoeten et al., 1974), and the electron dense material contains viral non-stuctural proteins (Wellink et al., 1988). The replication of viral RNA is closely associated with the membranes of the vesicles (Assink et al., 1973; De Zoeten et al., 1974). The vesicles probably are induced by the RNA1-encoded 60K protein and derived from the endoplasmic reticulum (Van Bokhoven et al., 1992; Carette et al., 2000). Virions assemble in the cytoplasm, and accumulate scattered and in clusters throughout the cytoplasm. They do not form crystalline arrays. They are also found within tubules which are involved in cell-to-cell transport, and penetrate the cell walls presumably through modified plasmodesmata (Fig. 8, Van Lent et al., 1990). The tubules contain the viral movement protein and are also formed on the surface of protoplasts (Fig. 9, Fig. 10, Fig. 11, Van Lent et al., 1991).
Ecology and Control
Because of the small plots used for cowpea growing and the extreme prevalence of the beetle vector, the use of insecticides for vector control is not practicable. Use of resistant cultivars offers the best means of disease control. Cowpea lines with resistance to multiple viruses including CPMV have been developed at the International Institute of Tropical Agriculture (Singh & Ntare, 1985). Some cowpea varieties like Tvu 470 and Arlington show immunity that is governed by a single dominant locus. However, protoplasts isolated from these varieties showed normal or only reduced accumulation of CPMV (Ponz et al., 1988; Saayer-Riep & De Jager, 1988). Other varieties like Tvu 1948 show immunity that is governed by a single recessive locus and show complete resistance to CPMV at the protoplast level (Patel, 1982).
Notes
The cowpea mosaic virus described here is the type species of the genus Comovirus (Wellink et al., 2000). Many viruses from other taxa have been isolated from Vigna spp. showing mosaic symptoms. The most notable of these are: cowpea aphid-borne mosaic virus (Bock & Conti, 1974); cowpea chlorotic mottle virus (Bancroft, 1971); cowpea mild mottle virus (Brunt & Kenten, 1974); sunnhemp mosaic virus (Kassanis & Varma, 1975); a strain of southern bean mosaic virus (Shepherd, 1971); a strain of bean common mosaic virus (Sachchidananda et al., 1973). Cowpea mosaic virus may be distinguished from these and other viruses of cowpea by the morphology of its particles and their behaviour in the ultracentrifuge and by its antigenic properties.
Figures

Chlorotic lesions in inoculated primary leaf of cowpea cv California Blackeye.

Systemic symptoms in cowpea cv California Blackeye.

Symptoms on a cowpea pod.

Yellow local lesions in inoculated Chenopodium amaranticolor leaf.
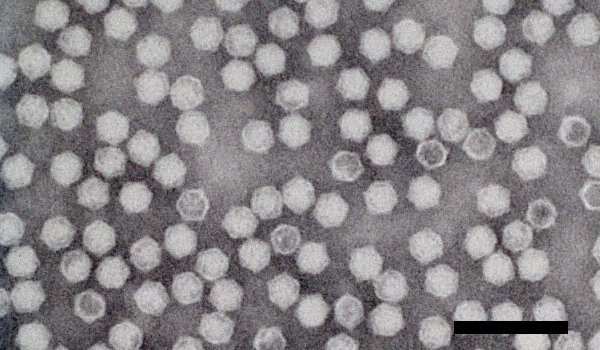
Electron micrograph of CPMV particles negatively stained with 2% PTA pH 6.8. Bar represents 100 nm.
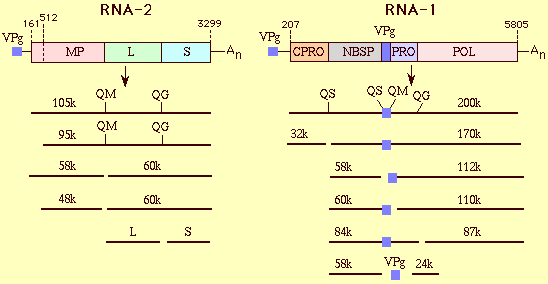
Organization and expression of the CPMV genome. The positions of the start and stop codons are indicated on the RNA, and functions of the proteins are indicated in the open reading frame of the RNA represented by an open bar. RNA2 is translated into two polyproteins due to initiation of translation at both positions 161 and 512. MP, movement protein; L, large capsid protein; S, small capsid protein; CPRO, processing regulator: NBSP, protein with nucleotide binding site; VPg, viral protein genome-linked; PRO, proteinase; POL, RNA-dependent RNA polymerase. Proteins are represented by single lines. The two amino acids forming the proteolytic cleavage sites are indicated on the polyproteins. Q, glutamine; G, glycine; S, serine; M, methionine. All intermediate and final cleavage products have been detected in infected cells.

Electron micrograph of a cytopathological structure in CPMV-infected Nicotiana benthamiana consisting of electron-dense structures (Eds) and small membranous vesicles (Ve). Ch, chloroplast; Va, vacuole. Bar represents 200 nm.
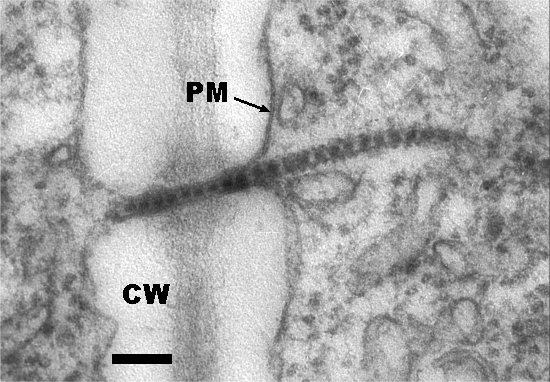
Electron micrograph of a tubular structure containing virus particles penetrating a cell wall (CW) through a presumably modified plasmodesma. PM, plasma membrane. Bar represents 100 nm.

Immunofluorescent image of a CPMV-infected cowpea protoplast treated with antiserum against the movement protein showing long tubular structures emerging from the protoplast surface. Bar represents 10 µm.

Electron micrograph of a thin section of a CPMV-infected protoplast showing a tubular structure. PM, plasma membrane; Cy, cytoplasm. Bar represents 100 nm.
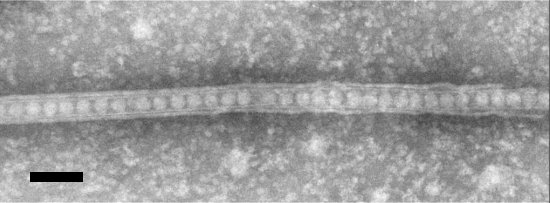
Electron micrograph of a negatively stained sample of a CPMV-infected protoplast showing a tubular structure. Bar represents 100 nm.
References list for DPV: Cowpea mosaic virus (378)
- Agrawal, Mededelingen van de LandbouwHogeschool Wageningen 64 (5): 53 pp., 1964.
- Agrawal & Maat, Nature 202: 674, 1964.
- Altmann & Lomonossoff, Journal of General Virology 81: 1111, 2000.
- Assink, Swaans & Van Kammen, Virology 53: 384, 1973.
- Bancroft, CMI/AAB Descriptions of Plant Viruses 49, 1971.
- Bliss & Robertson, Crop Science 11: 258, 1971.
- Bock, East African Agriculture and Forestry Journal 37: 60, 1971.
- Bock & Conti, CMI/AAB Descriptions of Plant Viruses 134, 1974.
- Bruening, Virology 37: 577, 1969.
- Bruening & Agrawal, Virology 32: 306, 1967.
- Brunt & Kenten, CMI/AAB Descriptions of Plant Viruses 140, 1974.
- Carette, Stuiver, Van Lent, Wellink & Van Kammen, Journal of Virology 74: 6556, 2000.
- Chant, Annals of Applied Biology 47: 565, 1959.
- Chant, Empire Journal of Experimental Agriculture 28: 114, 1960.
- Daubert, Bruening & Najarian, European Journal of Biochemistry 92: 45, 1978.
- De Jager, Abstracts, 2nd International Congress of Plant Pathology, no. 257, 1973.
- De Jager, Virology 70: 151, 1976.
- De Jager & Van Kammen, Virology 41: 281, 1970.
- De Zoeten, Assink & Van Kammen, Virology 59: 341, 1974.
- El Manna & Bruening, Virology 56: 198, 1973.
- Gamez, Turrialba 22: 249, 1972.
- Geelen, Rezelman & Van Kammen, Virology 51: 279, 1973.
- Gergerich & Scott, in The Plant Viruses 5, p.77, ed. B.D. Harrison & A.F. Murant, New York: Plenum Press, 1996.
- Gilmer, Whitney & Williams, in Proceedings of the 1st IITA Grain Legume Improvement Workshop, p. 269, Ibadan: IITA, 1974.
- Goldbach & Wellink, in The Plant Viruses 5, p.35, ed. B.D. Harrison & A.F. Murant, New York: Plenum Press, 1996.
- Jansen & Staples, Journal of Economic Entomology 64: 365, 1971.
- Jones & Barker, Annals of Applied Biology 83: 231, 1976.
- Kaiser, Danesh, Okhovat & Mossamebi, Iran Journal of Plant Pathology 4: 2, 1968.
- Kassanis & Varma, CMI/AAB Descriptions of Plant Viruses 153, 1975.
- Klootwijk, Klein, Zabel & Van Kammen, Cell 11: 73, 1977.
- Lin, Chen, Usha, Stauffacher, Dai, Schmidt, & Johnson, Virology 265: 20, 1999.
- Lomonossoff & Shanks, EMBO Journal 2: 2253, 1983.
- Niblett & Semancik, Virology 38: 685, 1969.
- Nickerson & Lane, Virology 81: 455, 1977.
- Partridge, Shannon, Gumpf & Colbaugh, Nature 247: 391, 1974.
- Patel, Phytopathology 72: 460, 1982.
- Ponz, Glascock & Bruening, Molecular Plant-Microbe Interactions 1: 25, 1988.
- Porta, Wang, Cheng, Chen, Baker & Johnson, Virology 204: 777, 1994.
- Saayer-Riep & De Jager, Netherlands Journal of Plant Pathology 94: 253, 1988.
- Sachchidananda, Singh, Nam & Verma, Zeitung für Pflanzenkrankheiten und Pflanzenschutz 80: 88, 1973.
- Semancik, Virology 30: 689, 1966.
- Shepherd, CMI/AAB Descriptions of Plant Viruses 57, 1971.
- Singh & Ntare, in Cowpea research, production and utilization, p105, ed. S.R. Singh and K.O. Rachie, Chichester: John Wiley & Sons, 1985.
- Stanley, Rottier, Davies, Zabel & Van Kammen, Nucleic Acids Research 5: 4505, 1978.
- Swaans & Van Kammen, Netherlands Journal of Plant Pathology 79: 257, 1973.
- Talens, Philippines Journal of Crop Science 4: 37, 1979.
- Taylor, Spall, Butler & Lomonossoff, Virology 255: 129, 1999.
- Thottappilly, Journal of Phytopathology 134: 265, 1992.
- Thottappilly & Rossel, in Cowpea research, production and utilization, p155, ed. S.R. Singh and K.O. Rachie, Chichester: John Wiley & Sons, 1985.
- Thottappilly & Rossel, Tropical Grain Legume Bulletin 34: 27, 1988.
- Valverde & Fulton, in The Plant Viruses 5, p.17, ed. B.D. Harrison & A.F. Murant, New York: Plenum Press, 1996.
- Van Bokhoven, Van Lent, Custers, Vlak, Wellink & Van Kammen, Journal of General Virology 73; 2775, 1992.
- Van Hoof, Surinaamse Landbouw 11: 131, 1963.
- Van Kammen, Virology 31: 633, 1967.
- Van Kammen, Virology 34: 312, 1968.
- Van Lent, Wellink & Goldbach, Journal of General Virology 71: 219, 1990.
- Van Lent, Storms, Van Der Meer, Wellink & Goldbach, Journal of General Virology 72: 2615, 1991.
- Van Wezenbeek, Verver, Harmsen, Vos & Van Kammen, EMBO Journal 2: 941, 1983.
- Virudachalam, Harrington, Johnson & Markley, Virology 141: 43, 1985.
- Wellink, Van Lent & Goldbach, Journal of General Virology 69: 951, 1988.
- Wellink, Le Gall, Sanfacon, Ikegami & Jones, in Virus Taxonomy 7, p. 691, ed. M.H.V. van Regenmortel et al., San Diego: Academic Press, 2000.
- Whitney & Gilmer, Annals of Applied Biology 77: 17, 1974.