Details of DPV and References
DPV NO: 383 December 2001
Family: Betaflexiviridae
Genus: Vitivirus
Species: Grapevine virus A | Acronym: GVA
Grapevine virus A
G. P. Martelli Dipartimento di Protezione delle Piante, Università degli Studi and Centro di Studio del CNR sui Virus e le Virosi delle Colture Mediterranee, 70126 Bari, Italy
M. Conti Istituto di Fitovirologia Applicata del CNR, 10135 Torino, Italy
A. Minafra Dipartimento di Protezione delle Piante, Università degli Studi and Centro di Studio del CNR sui Virus e le Virosi delle Colture Mediterranee, 70126 Bari, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Conti et al. (1980).
For disease description see Graniti & Martelli (1965); Martelli (1993).
Synonyms
A virus with filamentous particles about 800 nm long showing distinct transverse cross banding, occurring world-wide in Vitis species. Virus particles contain a single species of linear positive sense ssRNA of about 7.3 kb and a single protein species of Mr c. 22,000. The virus is a pathogen of grapevine from which it can be transmitted with difficulty by inoculation of sap to a very narrow range of herbaceous hosts. It is spread by propagation of infected material over medium and long distances and between plants by mealy bug vectors.
Main Diseases
The virus is the agent of Kober stem grooving (Digiaro et al., 1994; Garau et al., 1994; Chevalier et al., 1995) one of the components of the rugose wood complex (Savino et al., 1989). Rugose wood is a very severe disease of grafted vines, characterized by pits and grooves on the woody cylinder of the scion, the rootstock, or both (Graniti & Martelli, 1965) (Fig. 1). The virus induces distinct longitudinal grooves on the stem of the American rootstock hybrid Kober 5BB (Fig. 2) following graft-inoculation from grapevines (Garau et al., 1994), or approach grafting from Nicotiana spp. (B. Walter, personal communication). It can infect symptomlessly grapevine cultivars and rootstocks (Garau et al., 1991). The virus is responsible for crop losses from 5 to 22% in wine grape cultivars in Italy (Garau et al., 1997), and for decline and death of table grapevines affected also by leafroll disease (Digiaro et al., 1997).
Geographical Distribution
Probably occurs wherever Vitis vinifera is grown. Reported from Europe, the Mediterranean basin, Middle East, South Africa, China, Australia, North and Latin America (summarized by Boscia et al., 1997a).
Host Range and Symptomatology
Natural infection has been detected only in Vitis, frequently in association with other viruses (Conti & Milne, 1985; Digiaro et al., 1994). The experimental host range is restricted to a few Nicotiana species (Conti et al., 1980; Rosciglione et al., 1983; Monette & James, 1990a; Garau et al., 1994; Martelli et al., 1994). An isolate from Yemen induces local lesions in Chenopodium quinoa and C. amaranticolor and infects Datura stramonium symptomlessly (Martelli et al., 1994). The same isolate and one from Italy (Garau et al., 1995) induces local lesions in Gomphrena globosa. Virus isolates are readily transmitted between herbaceous hosts by mechanical inoculation of sap. Transmission from grapevine is erratic, especially if tissues from field-grown plants are used as inoculum. In vitro-grown grapevine explants and total nucleic acid extracts are better sources for mechanical transmission (Monette & James, 1990b; Martelli et al., 1994). For transmission, tissues are ground in 0.1M phosphate buffer containing 2.5% nicotine.
Diagnostic species
Chenopodium quinoa and C. amaranticolor. Chlorotic/necrotic local lesions in 8-10 days. No systemic invasion.
Gomphrena globosa. Chlorotic/necrotic local lesions in 7-8 days, sometimes followed by systemic interveinal mottling in 10-15 days.
Propagation species
Assay species
Strains
Two biological variants of the virus identified in Canada are differentiated by the severity of symptoms in N. benthamiana, but not serologically (Monette & James, 1990a). Serological variants exist, as reported from Switzerland (Gugerli et al., 1991), and as suggested by the weaker reaction consistently given by a French virus isolate with a monoclonal antibody to an Italian isolate (Boscia et al., 1992).
Transmission by Vectors
The virus is transmitted by the pseudococcid mealy bugs Pseudococcus longispinus, Ps. affinis, Planococcus ficus, Pl. citri (Rosciglione et al., 1983; Rosciglione & Castellano, 1985; Engelbrecht & Kasdorf, 1985; 1990a; 1990b; Garau et al., 1995) and the coccid mealy bug Neopulvinaria innumerabilis (Fortusini et al., 1997). Transmission by Ps . longispinus and Pl. ficus is semi-persistent. The virus is acquired after feeding for a minimum of 15 min and an optimum of 1-3 h, is retained for up to 48 h while fasting and up to 15 h while feeding, and is transmitted to plants with no latent period in 30 min to 3 h; transmissibility is lost after moulting. The virus is retained in the anterior part of the alimentary canal of Ps. longispinus (La Notte et al., 1997a and unpublished data). The virus is not transmitted by the aphids Myzus persicae and Macrosiphon euphorbiae (Conti et al., 1980).
Transmission through Seed
The virus was not seed-transmitted in N. clevelandii (Conti et al., 1980).
Transmission by Grafting
The virus is readily transmitted from vine to vine by grafting, which represents one of the major mechanisms for dissemination. The specific indicator Kober 5BB develops typical stem grooving following cleft-, machine-, or green-grafting (Martelli, 1993).
Serology
The virus is moderately immunogenic. Antisera with titres up to 1/512 were raised in rabbits and produced a single precipitin line in gel double diffusion tests, flocculent precipitates in microprecipitin tests, and decorated virus particles uniformly in immuno-electron microscopy tests (Conti et al., 1980; Castrovilli & Gallitelli, 1985; Engelbrecht & Kasdorf, 1990a) (Fig. 5). Four mouse monoclonal antibodies to purified virus particles have been produced with titres between 1/8,000 and 1/32,000, only one of which, derived from a surface antigenic determinant, decorated the whole virus particle in immuno-electron microscopy (Boscia et al., 1992). Cortical scrapings from mature canes are the best antigen source for serological testing (Boscia et al., 1997b). DAS-ELISA with polyclonal antisera can be used for virus detection in infected vines, but the uneven distribution and low concentration of the antigen in the host tissues affects the reliability of this test (summarized by Boscia et al., 1997b). More reliable results are obtained when polyclonal antisera are used for antigen trapping in protein-A pre-coated plates, followed by enzyme-conjugated monoclonal antibodies for antigen detection (Boscia et al., 1992). Monoclonal antibodies detect the virus in N. benthamiana using tissue blot assays (Choueiri et al., 1997). Antiserum raised to the recombinant virus movement protein expressed in Escherichia coli was used for sensitive virus detection in infected grapevine tissues (Rubinson et al., 1997) and for the identification of the intracellular sites of accumulation of the movement protein (Saldarelli et al., 2000).
Nucleic Acid Hybridization
A molecular probe obtained by random priming was used for virus detection in grapevine leaf extracts by spot hybridization (Gallitelli et al., 1985). Radioactive cDNA probes (Minafra et al., 1992) and digoxigenin-labelled riboprobes (Saldarelli et al., 1994) were used successfully for virus identification. Primers designed for the specific amplification of viral sequences detected virus in viruliferous mealy bugs and grapevine tissues by standard RT-PCR (Minafra & Hadidi, 1994), immuno-capture RT-PCR (Chevalier et al., 1995), or a spot-RT-PCR technique that minimized sample manipulation (La Notte et al., 1997b). Immuno-capture RT-PCR was estimated to be 1000-fold more sensitive than DAS-ELISA for virus detection in grapevines (Chevalier et al., 1995).
Relationships
The virus is the type species of the genus Vitivirus. (Martelli et al., 1997). It is serologically distantly related to other species in the same genus, i.e. Grapevine virus D (GVD), with which it shares a single antigenic determinant (Choueiri et al., 1997), Grapevine virus B (GVB) (Goszczynki et al., 1996), and Heracleum latent virus (HLV) (Murant et al., 1985). The replication-associated proteins contain signature sequences homologous to those of the genera Capillovirus, Carlavirus, Potexvirus, Trichovirus, and Tymovirus. The capsid protein has homology with that of capilloviruses and trichoviruses and is somewhat more distantly related to that of carlaviruses and potexviruses (Minafra et al., 1994, 1997).
Stability in Sap
In sap of N. clevelandii, the virus lost infectivity when heated for 10 min at 50 °C, diluted to 10-5, and stored at 20 °C for 6 days or at 5 °C for 15 days (Conti et al., 1980).
Purification
Glasshouse-grown plants of Nicotiana spp. (Conti et al., 1980; Castrovilli & Gallitelli, 1985; Engelbrecht & Kasdorf, 1987; Martelli et al., 1994) and in vitro cultures from N. benthamiana nodes (Monette & James, 1990a, 1990b) or Agrobacterium rhizogenes- transformed N. benthamiana roots (Lupo et al., 1994) have been used as source material for virus purification. The following purification methods have been used successfully.
1. Grind leaves of N. clevelandii at 4 ºC with 3 vol of 0.05M tris-HCl buffer pH 7.8, containing 0.01M MgSO4 (TM buffer). Filter the homogenate through nylon cloth and centrifuge at 4,000 rpm for 10 min, then at 8,000 rpm for an additional 10 min. Clarify by adding 3.5 ml bentonite suspension to every 100 ml and centrifuge at 10,000 rpm for 10 min. Add 5 g of PEG (mol. wt 6,000) and 0.5 ml of 5M NaCl to each 100 ml of the supernatant fluid, place on ice for 2-3 h, then centrifuge at 10,000 rpm for 15 min. Resuspend the pellets in 10 ml of TM buffer and centrifuge at 10,000 rpm for 10 min. Layer the supernatant fluid onto a pre-formed linear gradient of Cs2SO4 in 0.01M phosphate buffer pH 7.5 and centrifuge at 37,500 rpm in a Beckman SW41 rotor. Extract the virus-containing band(s) and dialyze against 0.05M phosphate buffer, pH 7.5 or concentrate by high speed centrifugation (Conti et al., 1980).
2. Grind tissues of N. benthamiana in liquid nitrogen, transfer to a blender with 3 vol of TM buffer and homogenize for 20 sec. Filter the homogenate through cheesecloth and centrifuge at 3,300 g for 10 min. To the supernatant fluid add Triton X-100 to 2% and stir on ice for 45 min. Add NaCl to 0.2M, PEG-6,000 to 0.6%, 2-mercaptoethanol to 0.2%, and stir the mixture for 1.5 h. Centrifuge at 12,000 g for 20 min and resuspend the pellets in 10 ml of TM buffer and centrifuge for 10 min at 5,000 g. Layer 3 ml of the supernatant fluid on top of a step gradient consisting of 4 ml each of 20, 30, 40, and 50% sucrose in TM buffer, and centrifuge at 18,000 rpm for 3.5 h in a SW.25 Beckman rotor. Collect the virus-containing band, concentrate the virus by high speed centrifugation and resuspend the pellets in 3.5 ml Tris-HCl pH 7.2, containing 0.01M MgCl2. Add 0.53 g of Cs2SO4 and dissolve by vortexing. Layer this preparation onto a 1 ml pad of 53% Cs2SO4 in 0.1M Tris-HCl pH 8.2 and 0.01M MgCl2 and centrifuge for 18 h at 24,000 rpm in a Beckman SW 50.1 rotor. Recover the virus-containing band, dialyze overnight against TM buffer and then pellet the suspension through a 1 ml cushion of 20% sucrose in TM buffer by centrifuging for 4 h at 15,000 rpm in a SW 50.1 rotor (Monette & James, 1990b).
3. Grind tissues of N. benthamiana in 3 vol of 0.01M Tris-HCl buffer pH 7.6 (extraction buffer), filter the homogenate through cheesecloth and clarify by adding 4% Mg++-activated bentonite while stirring. Centrifuge for 15 min at 10,000 g, add 8% PEG (mol. wt 6,000) and 1% NaCl and allow it to stand in the cold for 1 h. Centrifuge at 6,000g for 10 min, resuspend the pellet in extraction buffer overnight and centrifuge at 89,000 g for 1 h. Resuspend the pellets overnight in a small volume of extraction buffer, clarify by centrifuging at 6,000 g for 10 min and then layer the supernatant fluid on 35% sucrose columns that were frozen and then thawed before use and centrifuge at 24,000 rpm in a Beckman SW 27 rotor. Recover the virus-containing band and dialyze it against extraction buffer (Martelli et al., 1994).
All these methods usually yield a single virus-containing band in sucrose density gradients and which is sharper when centrifuged through Cs2SO4 gradients. Castrovilli & Gallitelli (1985) reported the occasional presence of two centrifugal components in Cs2SO4 gradients the upper of which was about ten-fold less infective that the faster sedimenting component. Yields are up to 1 mg virus/100g of tissue (Castrovilli & Gallitelli, 1985).
Properties of Particles
Sedimentation coefficient (s20,w) not extrapolated to infinite dilution: 92 ± 2S (Castrovilli & Gallitelli, 1985).
A260/A280 : 1.52; Amax (260nm)/Amin (243 nm) : 1.12 (Boccardo & d'Aquilio, 1981).
Particle Structure
The virus particles are flexuous filaments c. 800 x 12 nm in length, showing distinct cross banding (Fig. 6), helically constructed with a pitch of 3.3 nm and about 10 subunits per turn of the helix (Conti et al., 1980).
Particle Composition
Nucleic acid. Virus particles contain a single species of linear, positive sense, ssRNA that accounts for about 5% of the particle weight. The RNA is completely denatured at 75 °C. It has Amax/Amin ratio of 1.82-1.86, A260/A280 ratio of 1.72-1.79, depending on the isolation method used. It has the following base composition: U, 22.3; C, 24.9; A, 24.4; G, 28.4 mol % (Boccardo & d'Aquilio, 1981). Regardless of the method of isolation (Boccardo & d'Aquilio,1981; Castrovilli & Gallitelli, 1985) the RNA is infective, and induces typical virus symptoms when manually inoculated to N. clevelandii (Boccardo & d'Aquilio, 1981).
Proteins. The virus coat protein accounts for about 95% of the particle weight and is composed of a single polypeptide of Mr 22,0000 - 27,000, as calculated by polyacrylamide gel electrophoresis. The most frequently reported value is 22,000-22,500, which is in very good agreement with the value of 21,600 deduced from the amino acid sequence of the coat protein gene (summarized by Boscia et al., 1997a). The coat protein does not contain tryptophan, as shown by spectrophotometric analysis (Boccardo & d'Aquilio, 1981), and sequencing (Minafra et al., 1994). Virus coat protein transgenically expressed in N. benthamiana plants was able to encapsidate the genomic RNA of Grapevine virus B, following inoculation with the latter virus (Buzkan et al., 2001).
Genome Properties
The viral genome has been totally sequenced and contains 7,349 nt (accession No. X75433). It is capped at the 5' terminus and polyadenylated at the 3' terminus, and contains five slightly overlapping ORFs (Minafra et al., 1994, 1997) (Fig. 7). The 5' region initiates with an A/T rich (60%) untranslated sequence (UTR) of 86 nt, and the 3' region terminates with an UTR of 68 nt, not including the poly(A) tail. ORF 1 (nt 87 to 5210) encodes a polypeptide of 195 kDa showing extensive homologies with the replication-related proteins of RNA viruses, i.e. methyltransferase domain in the N-terminal region (aa 67-245), helicase motifs (aa 922-1145) in the central region, and the RNA dependent RNA polymerase domain (aa 1395-1623) in the C- terminal region of the molecule. ORF2 (nt 5179-5712), which partially overlaps ORF1 and ORF3, encodes a polypeptide of 19.8 kDa with no significant homology with proteins in databases. ORF3 (nt 5654-6490) codes for a polypeptide of 31 kDa with the conserved motifs of movement proteins of the 30K `superfamily'. ORF4 (nt 6414-7010) encodes the coat protein cistron of 21.6 kDa. ORF 5 (nt 7015-7281) codes for a polypeptide of 10.1 kDa with putative nucleotide binding activity (Minafra et al., 1994, 1997). The 19.8 kDa product is transiently expressed in N. benthamiana (P. Saldarelli, personal communication). In the same host, the movement protein is transiently expressed in the cell membranes but accumulates in the cytosol and in the cell walls in association with plasmodesmata (Saldarelli et al., 2000). The expression strategy, as suggested by the dsRNA pattern from infected tissues, is based on direct translation from genomic RNA of the 5' proximal ORF and subgenomic RNA production for the translation of downstream ORFs. Viral dsRNAs from infected plants consist of four major bands with sizes of 7.6, 6.48, 5.68, and 5.1 kbp, respectively (Boscia et al., 1994). Virus replication is in the cytoplasm, possibly in association with membranous vesicles.
Satellite
No accessory RNA molecules have been detected (Boccardo & d'Aquilio, 1981; Castrovilli & Gallitelli, 1985).
Relations with Cells and Tissues
Virus isolates of different geographical origin induce little ultrastructural modifications in parenchyma cells of several Nicotiana species, G. globosa, and Vitis vinifera. The cell architecture is preserved, major organelles have a normal appearance, and virus particles are few and form small, scattered aggregates (Rosciglione et al., 1983; Faoro et al., 1991; Monette & Godkin, 1992; M. A. Castellano, personal communication). By contrast, vascular bundle cells (sieve tubes, companion and vein border cells) show a much deranged cytology, extended cell wall thickening originated from deposits of callose-like substances, proliferation of endoplasmic membranes, vesicular evaginations of the tonoplast containing finely fibrillar material resembling dsRNA. Virus particle aggregates, in the form of bundles, whorls, banded bodies, or stacked layers (Fig. 8), are massive, sometimes protruding into the vacuole from tonoplast evaginations, or filling most of the cell lumen (Rosciglione et al., 1983, Monette & Godkin, 1992; Lupo et al., 1994; M. A. Castellano, personal communication). Intracelluar virus particles were distinguished from P-protein filaments by differential fixation (Faoro & Tornaghi, 1991) and by immunogold labeling from closterovirus particles in doubly infected cells (Faoro et al., 1991). In N. benthamiana, the movement protein is transiently expressed in the cell membranes, and accumulates in the cell wall at the level of plasmodesmata and in the cytosol, as detected by immunogold labeling (Saldarelli et al., 2000).
Ecology and Control
The inoculum potential in the field seems to be fairly high, as the virus was consistently detected in over 70% of the mealy bug populations collected in rugose wood-infected vineyards from different European and Mediterranean countries (La Notte et al., 1997b). The vectors are not easy to control by pesticides as they overwinter under the bark of the vines and possess an unwettable waxy covering. Thus, local virus spread is difficult to restrain. By contrast, long distance spread can be controlled readily by the production and distribution of healthy propagative material (budwood, rooted rootstocks, or grafted vines). The virus can be eliminated by heat therapy (Chevalier et al., 1995), meristem tip culture, or a combination of the two, with fairly high efficiency (up to 85%) (G. Bottalico & V. Savino, personal communication). No natural sources of resistance to the virus are known but the possibility of using pathogen-derived resistance in Vitis is being explored. Using a N. benthamiana model system, several virus resistant plant lines were obtained by transformation with the coat protein and the movement protein genes of the virus. Transgene expression was detected in these plants and in transformed grapevine explants (Minafra et al., 1998; Saldarelli et al., 2000).
Notes
The virus was first classified as a member of the Closterovirus group (Bar-Joseph & Murant, 1982), then as a tentative species in the genus Trichovirus (Martelli et al., 1994), and finally to the genus Vitivirus, as the type species. The genus Vitivirus was separated from the genus Trichovirus because of differences in genome structure and organization, and in the biological and epidemiological behaviour of their respective species (Martelli et al., 1997). The virus is very similar to Grapevine virus D, with which it shares a cryptotope and a similar dsRNA electrophoretic pattern (Abou-Ghanem et al., 1997). However, whether these two viruses are each involved in the aetiology of the same disease (Kober stem grooving) remains to be determined.
Acknowledgements
We thank R. G. Milne and M. A. Castellano for the micrographs.
Figures

Rugose wood symptoms at the base of an infected vine. Note the pitting of exposed wood.

Grooving on the woody cylinder of an infected Kober 5BB rootstock.

Stunting and systemic mottling in infected N. benthamiana. Healthy plant on the right.

Vein clearing and deformation of systemically infected leaves of N. clevelandii.

Virus particle decorated by virus-specific antibodies conjugated with colloidal gold. Bar = 100 nm (courtesy of R. G. Milne).

Virus particles mounted in uranyl acetate. Bar = 100 nm (courtesy of R. G. Milne).
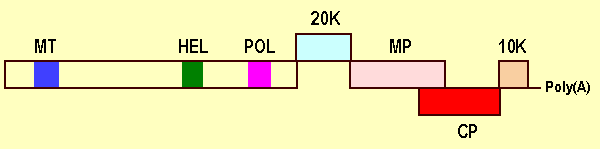
Virus genome organization showing the relative positions of the ORFs and their translation products. MT, methyltransferase; HEL, helicase; POL, polymerase; MP, putative movement protein; CP, coat protein.

Accumulations of virus particles in the cytoplasm of a differentiating sieve tube of N. benthamiana. Bar = 250 nm (courtesy of M. A. Castellano).
References list for DPV: Grapevine virus A (383)
- Abou-Ghanem, Saldarelli, Minafra, Buzkan, Castellano & Martelli, Journal of Plant Pathology 78: 15, 1997.
- Bar-Joseph & Murant, CMI/AAB Descriptions of Plant Viruses 260, 1982.
- Boccardo & d'Aquilio, Journal of General Virology 53: 181, 1981.
- Boscia, Aslouj, Elicio, Savino, Castellano & Martelli, Archives of Virology 127: 185, 1992.
- Boscia, Abou-Ghanem, Saldarelli, Minafra, Castellano, Garau, Savino & Martelli, Rivista di Patologia Vegetale 3 (S.V): 11, 1994.
- Boscia, Minafra & Martelli, in Filamentous Viruses of Woody Crops, p. 19, ed. P. L. Monette, Trivandrum: Research Signpost, 1997a.
- Boscia et al., in Sanitary Selection of the Grapevine. Protocols for the Detection of Viruses and Virus-like Diseases, p. 129, ed. B. Walter, Les Colloques INRA N° 86, Paris: INRA Editions, 1997b.
- Buzkan, Minafra, Saldarelli, Castellano, Dell'Orco, Martelli, Gölles & Laimer da Camara Machado, Journal of Plant Pathology 83: 37, 2001.
- Castrovilli & Gallitelli, Phytopathologia Mediterranea 24: 219, 1985.
- Chevalier, Greif, Clauzel, Walter & Fritsch, Journal of Phytopathology 143: 369, 1995.
- Choueiri, Abou-Ghanem & Boscia, Vitis 36: 39, 1997.
- Conti & Milne, Phytopathologia Mediterranea 24: 110, 1985.
- Conti, Milne, Luisoni & Boccardo, Phytopathology 70: 394, 1980.
- Digiaro, Boscia, Simeone & Savino, Extended Abstracts 12th Meeting of ICVG, Lisbon, 1997: 169, 1997.
- Digiaro, Popovic Bedzrob, D'Onghia, Boscia & Savino, Phytopathologia Mediterranea 33: 187, 1994.
- Engelbrecht & Kasdorf, Phytopathologia Mediterranea 24: 101, 1985.
- Engelbrecht & Kasdorf, South African Journal of Enology and Viticulture 8: 23, 1987.
- Engelbrecht & Kasdorf, Phytophylactica 22: 341, 1990a.
- Engelbrecht & Kasdorf, Phytophylactica 22: 347, 1990b.
- Faoro & Tornaghi, Giornale Botanico Italiano 125: 998, 1991.
- Faoro, Tornaghi & Belli, Journal of Phytopathology 133: 297, 1991.
- Fortusini, Scattini, Prati, Cinquanta & Belli, Extended Abstracts 12th Meeting of ICVG, Lisbon, 1997: 121, 1997.
- Gallitelli, Savino & Martelli, Phytopathologia Mediterranea 24: 221, 1985.
- Garau, Prota & Prota, Proceedings of the 10th Meeting of ICVG, Volos 1990: 175, 1991.
- Garau, Prota, Piredda, Boscia & Prota, Vitis 33: 161, 1994.
- Garau, Prota, Boscia, Fiori & Prota, Vitis 34: 67, 1995.
- Garau, Fiori, Prota, Tolu, Fiori & Prota, Extended Abstracts 12th Meeting of ICVG, Lisbon, 1997 : 171, 1997.
- Goszczynski, Kasdorf & Pietersen, Journal of Phytopathology 144: 11, 1996.
- Graniti & Martelli, Proceedings of the International Symposium on Virus and Vectors on Perennial Hosts with Special reference to Vitis, Davis 1965 : 168, 1965.
- Gugerli, Rosciglione, Brugger, Bonnard, Ramel & Tremea, Proceedings of the 10th Meeting of ICVG, Volos 1990: 59, 1991.
- La Notte, Minafra & Saldarelli, Journal of Virological Methods 66: 103, 1997a.
- La Notte, Buzkan, Choueiri, Minafra & Martelli, Journal of Plant Pathology 78: 79, 1997b.
- Lupo, Martelli, Castellano, Boscia & Savino, Plant Cell Tissue and Organ Culture 36: 291, 1994.
- Martelli (ed.), Graft-transmissible Diseases of Grapevine. Handbook for Detection and Diagnosis. Rome, FAO Publication Division, 263pp, 1993.
- Martelli, Candresse & Namba, Archives of Virology 134: 451, 1994.
- Martelli, Minafra & Saldarelli, Archives of Virology 142: 1929, 1997.
- Minafra & Hadidi, Journal of Virological Methods 47: 175: 1994.
- Minafra, Russo & Martelli, Vitis 31: 87, 1992.
- Minafra, Saldarelli, Grieco, & Martelli, Archives of Virology 137: 249, 1994.
- Minafra, Saldarelli & Martelli, Archives of Virology 142: 417, 1997.
- Minafra, Gölles, da Camara Machado, Saldarelli, Buzkan, Savino, Martelli, Katinger, & Laimer da Camara Machado, Journal of Plant Pathology 80: 197, 1998.
- Monette & Godkin, Canadian Journal of Plant Pathology 14: 1, 1992.
- Monette & James, Plant Disease 74: 898, 1990a.
- Monette & James, Plant Cell Tissue and Organ Culture 23: 131,1990b.
- Murant, Duncan & Roberts, Report of the Scottish Crop Research Institute 1984: 182, 1985.
- Rosciglione, Castellano, Martelli, Savino & Cannizzaro, Vitis 22: 331, 1983.
- Rosciglione & Castellano, Phytopathologia Mediterranea 24: 186, 1985.
- Rubinson, Galiakparov, Radian, Sela, Tanne & Gafny, Phytopathology 87: 1041, 1997.
- Saldarelli, Gugliemi Montano & Martelli, Vitis 33: 157, 1994.
- Saldarelli, Minafra, Castellano & Martelli, Archives of Virology 145: 1535, 2000.
- Savino, Boscia & Martelli, Proceedings of the 9th ICVG Meeting, Kyriat Anavim 1987: 91, 1989.