Details of DPV and References
DPV NO: 393 October 2002
Family: Potyviridae
Genus: Tritimovirus
Species: Wheat streak mosaic virus | Acronym: WSMV
This is a revised version of DPV 48
Wheat streak mosaic virus
Roy French USDA, ARS, Department of Plant Pathology, University of Nebraska, Lincoln, USA
Drake C. Stenger USDA, ARS, Department of Plant Pathology, University of Nebraska, Lincoln, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by McKinney (1937).
- Synonyms
- Wheat viruses 6 and 7 for severe and mild isolates of the virus, respectively (McKinney, 1937).
A mite-transmitted virus with filamentous particles (about 15 x 700 nm) with a monopartite, single-stranded RNA genome. The virus is readily transmitted by mechanical inoculation of plant sap to many species in the Poaceae but does not infect any dicotyledonous plants.
Main Diseases
Causes a severe mosaic disease in winter wheat (Fig. 1, Fig. 2) and less frequently in spring wheat. Plants are usually stunted and develop a rosette appearance. Tillers of infected plants tend to be less erect than those of uninfected plants. Symptoms are not usually apparent in autumn sown plants in early spring but become noticeable as temperatures rise above 10°C. Plants infected in the spring usually have a milder mosaic with reduced stunting. As WSMV-infected plants often develop sterile heads, the effects on yield can be severe. Also causes a mild mosaic in some varieties of maize, primarily in regions where maize and winter wheat crops overlap. The disease is most serious when wheat is infected in autumn and leads to severe stunting (Fig. 1), reduced tillering, poor seed set, and lower seed weight. Losses can be 100% in severely affected fields. It also causes mosaic disease in some millets (Brakke, 1987, Edwards & McMullen, 1987).
Geographical Distribution
Canada, the Czech Republic, Hungary, Iran, Jordan, Mexico, Poland, Rumania, Russia, Turkey, USA, Yugoslavia (Jezewska, 2000; Rabenstein et al., 2002; Sánchez-Sánchez et al. 2001; Slykhuis, 1967). The virus is common and of greatest economic importance in the Great Plains region of North America.
Host Range and Symptomatology
Infects most (see STRAINS) varieties of wheat (Triticum aestivum), oats (Avena sativa), barley (Hordeum vulgare), and rye (Secale cerale), and certain inbred lines or varieties of maize (Zea mays), sorghum (Sorghum vulgare), and millets (Panicum, Setaria, and Echinochloa spp.); all develop yellow-green mosaic. Infects many wild grass species in the genera Aeiglops, Agropyron, Bouteloua, Bromus, Cenchrus, Digitaria, Echinochloa, Elymus, Eragrostis, Haynaldia, Hordeum, Lolium, Panicum, Phalaris, Poa, Orizopsis, Setaria, and Stipa. Does not infect Agropyron repens, Bromus inermis, Hordeum jubatum, Oryza sativa (rice) or Saccharum officinarum (sugar cane).
No dicotyledonous hosts have been reported (Brakke, 1971; Kahn & Dickerson, 1957; Seifers, et al., 1996; Sill & Agusioba, 1953; Sill & Conin, 1953).
In the field, virus symptoms first appear as temperatures become warmer in mid to late spring. Infected plants exhibit mosaic symptoms with tillers much less erect than those of uninfected plants. Symptoms persist until plant maturity.
- Diagnostic species
Causes systemic mosaic symptoms, usually with stunting in nearly all varieties of wheat (Fig. 2), barley, and oats, and some maize varieties, at both cool and warm temperatures. Some isolates give sporadic necrotic lesions on some sorghum (Sorghum bicolor) varieties.
- Propagation species
Wheat. Essentially any winter wheat variety is suitable. Historically the variety Michigan Amber has been used because it resists mildew in typical greenhouse conditions.
- Assay species
No satisfactory local lesion assay host has been reported. Systemic assays of diluted sap may be made on wheat seedlings. Vector transmission tests also are best done with wheat, which is also a good host of the mite.
Strains
Many variants can be identified on the basis of symptom severity, but these differences are highly dependent on growing conditions and host varieties used. McKinney (1956) denoted a Type (ATCC PV57) and a mild (ATCC PV91) strain for cross protection experiments. Nucleotide sequences of isolates from USA, Eurasia, and Mexico differ by 10 to 20% (Choi et al., 2001, Rabenstein et al., 2002). Isolates also differ in serological properties (Montana et al., 1996). Isolates have been grouped by the electrophoretic mobilities of the virus coat protein in polyacrylamide gels and the ability to infect particular maize or sorghum lines (Brakke & Samson, 1981; Brakke et al., 1990; Seifers, et al., 1996). Certain European isolates do not infect oats or barley (Rabenstein et al., 2002).
Transmission by Vectors
Transmitted by an eriophyid mite, Aceria tosichella (Fig. 3). Earlier literature refers to this species (common name: wheat curl mite) as either A. tulipae or Eriophyes tulipae. Both adult and nymphal stages of the mite can transmit virus, but only nymphs may acquire virus. Mites tend not to settle down and feed upon transfer to new leaves, which makes timing studies difficult. The virus acquisition access feeding time is 15 min or more and the virus persists for at least 9 days in mites kept on virus-immune plants. There is no evidence either way regarding the presence of a latent period following virus acquisition. Virus inoculation access feeding time is several hours although an access feeding time of 15 min resulted in 2.5% transmission. The virus is not vertically transmitted to progeny through the egg. Mite transmission can serve as a bottleneck to reduce genetic variation in the virus (Amrine & Stansy, 1994; Hall et al., 2001; Orlob, 1966; Slykhuis, 1955; Staples & Allington, 1956).
Paliwal and Slykhuis (1967) detected WSMV by serology and by infectivity assays in homogenates of A. tosichella reared on WSMV-infected plants but not in homogenates of two non-vector mite species reared on similar plants. These observations led to speculation that back flow of gut contents during feeding or, excretion of intact virus, were potential routes for virus transmission by A. tosichella. Electron microscopy of thin sections of viruliferous A. tosichella detected aggregates of virus particles in the posterior midgut and hindgut that persisted for at least 5 days after feeding (Paliwal & Slykhuis, 1967). In later electron microscope studies, Paliwal (1980) reported the detection of WSMV particles in the haemocoel and salivary glands, but not in the hindgut, of viruliferous A. tosichella. When Abacarus hysterix, a non-vector of WSMV, reared on WSMV-infected plants, was examined by electron microscopy, small numbers of degraded WSMV particles were detected only in the gut. Although such studies may suggest that virus transmission by A. tosichella may be circulative, further evidence for this is lacking.
Transmission through Seed
The virus is seed transmitted in maize at 0.1-0.2% (Hill et al., 1974).
Serology
The virus is a comparatively good immunogen and antisera with titres of 1/256 to 1/1024 (microprecipitin assays) have been prepared by intramuscular injection of 1.2 mg purified virus mixed with Freund's complete adjuvant on days 1 and 8, an intravenous injection of 1.2 mg virus on day 11, followed by intramuscular injection with adjuvant on day 18 (Ball & Brakke, 1968). Early tests included microprecipitin assays, immunodiffusion with detergent disrupted virions, and leaf dip serological methods (Ball & Brakke, 1968). Sherwood (1987) found DAS-ELISA, Western blotting and filter paper binding assays to be suitable for virus detection. Sherwood et al., (1990) produced monoclonal antibodies that were subsequently shown to react with some but not all virus isolates (Montana et al., 1996). Virions in sap from young infected leaves are decorated with gold-labelled polyclonal antibody but not those from older leaves (Langenberg, 1989). Virus is easily detected in its mite vector using fluorescein-labelled antiserum (Mahmood et al., 1997).
Relationships
The virus is serologically related to Oat necrotic mottle virus and analysis of nucleotide sequences confirm a close relationship (75% identity) between these viruses (Rabenstein et al., 2002) and a more distant relationship (60% identity) with Brome streak mosaic virus (Stenger et al., 1998). The virus is the type species of the genus Tritimovirus, but was previously grouped with other mite-transmitted viruses in the genus Rymovirus. However, true members of the latter genus are quite divergent from tritimoviruses (Stenger et al., 1998).
Stability in Sap
In wheat sap the dilution end-point is 10-3 to 10-4 and the thermal inactivation point is 54°C (10 min). Infectivity in wheat sap survives for 4 but not 8 days at 20°C, and for 1 but not 2 months at 2°C. Leaves desiccated over CaCl2 and stored at 2°C retain infectivity for at least 16 years (McKinney et al., 1965).
Purification
A slight modification of the method described by Brakke (1971) gives reliable yields. Harvest the youngest 2 or 3 leaves of systemically infected wheat seedlings about 10 days after inoculation and triturate in 2 volumes (w/v) of 0.01 M K2HPO4. Adjust the pH to 6.1 with 1 M acetic acid and centrifuge for 30 min at 12,000 g. Remove and adjust the supernatant fluid to pH 8.0 and add 1/10 vol 0.1 M sodium citrate, pH 8.0 and bring to 1% Triton X-100. Layer the mixture over 4 ml cushion of 20% (w/v) sucrose in 0.01 M sodium citrate, pH 8.0 and centrifuge for 2 hr at 85,000 g. Resuspend the pellets in 0.01 M sodium citrate, pH 8.0 and clarify by centrifuging for 5 min at 3,000 g. Preparations can be purified further by centrifugation in sucrose (10-40%) density gradients (Brakke et al., 1990).
Alternative methods for purifying virus from frozen material (Slykhuis & Bell, 1965) and from infected maize leaves (Uyemoto & Ferguson, 1980) have been described.
Properties of Particles
Sedimentation coefficient as determined in sucrose density gradients is about 165S. Buoyant density in CsCl is 1.30-1.32 g.cm-3.
A260/280: 1.37; A230/260: 2.7; A260/320: 16 (Brakke & Van Pelt, 1968).
Particle Structure
Virions are flexuous rods c.15 nm in diameter and c.700 nm long (Fig. 4). Uranyl acetate and phosphotungstate are suitable stains for electron microscopy.
Particle Composition
Nucleic acid: Virus particles contain a single species of ssRNA. The complete nucleotide sequence (9,339 to 9,384 nt) of five isolates have been determined and deposited in GenBank as Accession Nos. AF057533, AF285169, AF285170, AF454454, AF454455 (Stenger et al., 1998; Choi et al., 2001; Rabenstein et al., 2002).
Protein: In polyacrylamide gel electrophoresis, virus particle preparations typically contain a major protein of Mr 45,000-47,000. Electrophoretic mobility is anomalous because coat protein size estimates vary with gel concentration and do not correspond to the molecular mass calculated from its predicted amino acid sequence. Most preparations contain varying amounts of smaller (Mr 31,000-40,000), partially degraded forms of the protein (Brakke et al., 1990). By analogy to other potyviruses, virions presumably contain a VPg protein linked to the 5'-end of the genomic RNA, but this has not been determined experimentally.
Genome Properties
The RNA molecule encodes a single ORF of about 350 kDa (Fig. 5) that is processed into mature proteins analogous to those of other potyviruses. P1 (41.2 kDa) has proteinase activity and probably cleaves at a GLRWY/G motif; HC-Pro (44.9 kDa) also is a proteinase; P3 (32.6 kDa) is of unknown function; CI (75.6 kDa) is the cylindrical inclusion body protein and has motifs shared by the DEAD/H class of RNA helicases; 6K1 and 6K2 are two short proteins of unknown function flanking CI; NIa (49.9 kDa) is the viral proteinase and probably also encodes a genome-linked VPg sequences; NIb (58.5 kDa) is the RNA-dependent RNA polymerase and is followed by the carboxy-terminal coat protein (40.9 kDa) (Stenger et al., 1998; Choi et al., 2000a; 2002).
Infectious cDNA clones have been produced for the virus. Foreign gene sequences have been inserted into these and expressed in wheat, barley, maize and oat (Choi et al., 1999; 2000b).
Relations with Cells and Tissues
Typical pinwheel inclusions are produced in infected plant cells and some are observed firmly attached to, and protruding through plasmadesma. The inclusions are made of CI protein but are associated with virions and viral coat protein. Aggregates of virus particles occur in the cytoplasm. Amorphous inclusions accumulate, often near deformed nuclei, and cytoplasm density increases. Proliferation of membranes and ribosomes is also observed. Cellular organelles become degraded late in infection (Gao & Nassuth, 1992; 1994; McMullen & Gardner, 1980; Langenberg, 1986; 1993; Shepard & Carroll, 1967).
Ecology and Control
Only juvenile mites can acquire and transmit WSMV but, once they acquire WSMV, they retain the ability to transmit virus as adults. Mites are dispersed solely by wind currents. Disease management relies on eliminating refuges of the virus and its mite vector during the period between summer harvest and autumn planting. This mainly involves control of volunteer wheat but limiting other potential hosts such as annual grassy weeds may also help. Delaying autumn planting until mite populations subside provides additional disease control. Chemical control of the vector is not economically feasible. No effective source of resistance to WSMV has been found in wheat but three loci for genetic resistance have been identified in maize. Host-plant resistance has been introgressed into wheat from wild relatives and may provide an additional means of control. Approaches to control using transgenic wheat expressing WSMV coat protein and NIa (replicase) for disease resistance also have been partially successful, and use of wheat varieties resistant to the mite vector may help reduce disease incidence. (Brakke, 1987; Chen et al., 1999; Christian & Willis, 1993; McMullen et al., 1994; Harvey et al., 1994; Seifers et al., 1995; Sivamani et al., 2000; 2002).
Notes
There are several viruses with similar morphology that infect wheat. However, of these, Agropyron mosaic virus infects Agropyron repens and Hordeum mosaic virus infects Hordeum jubatum, species that are immune to WSMV. Another virus, Wheat spindle streak mosaic virus is difficult to transmit mechanically and mosaic symptoms due to this virus fade as temperatures increase above 15°C, unlike those of WSMV. Brome streak mosaic virus can be distinguished by serologically from WSMV. The other tritimovirus, Oat necrotic mottle virus, does not infect wheat. Reverse transcription-polymerase chain reaction based assays are convenient for diagnosing WSMV (Hall et al., 2001).
In mixed infections with Maize chlorotic mottle virus titres of both viruses are increased several-fold (Scheets, 1998).
Figures

A severely stunted infected wheat plant with oblique growth habit of tillers.

Wheat leaves showing a range of chlorotic symptoms; bottom leaf is healthy.

Scanning electron micrograph of the wheat curl mite vector, Aceria tosichella. Bar represents 10 µm.

Transmission electron micrograph of virus particles stained with uranyl acetate. Bar represents 100 nm.
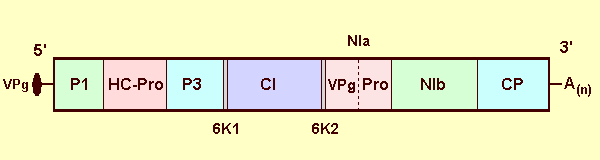
Genome organization; box represents the polyprotein open reading frame with vertical lines demarcating sites of cleavage by viral proteinases. Predicted mature proteins are labelled by analogy to other potyviruses: P1 - P1 protein, HC-Pro - helper component and proteinase (note that it is not known whether HC-Pro of WSMV is involved in transmission of the virus by mites), P3 - P3 protein, 6K1 and 6K2 - 6 kDa proteins, CI - cytoplasmic inclusion protein, NIa - comprised of a 5' genome-linked protein (VPg) and proteinase (Pro) domains, NIb - replicase, and CP - coat protein.
References list for DPV: Wheat streak mosaic virus (393)
- Amrine & Stansy, Catalogue of the Eriophyoidea (Aceria: Prostigmata) of the World, Indira Publishing, Bloomfield, MI, USA. 1186 pp., 1994.
- Ball & Brakke, Virology 36: 152, 1968.
- Brakke, C.M.I./A.A.B. Descriptions of Plant Viruses 48, 4 pp., 1971.
- Brakke, in, Wheat and Wheat Improvement, 2nd ed., ed E. G. Heyne, ASA-CSSA-SSSA, Madison WI, USA., 585, 1987.
- Brakke & Samson, Phytopathology 71: 823, 1981.
- Brakke, Skopp & Lane, Phytopathology 80: 1401, 1990.
- Brakke & Van Pelt, Analytical Biochemistry 26: 242, 1968.
- Chen, Conne, Laroche, Fedak & Thomas, Genome 42: 289, 1999.
- Choi, French, Hein & Stenger, Phytopathology 89: 1182, 1999.
- Choi, Stenger & French, Virology 267: 185, 2000a.
- Choi, Stenger, Morris & French, Plant Journal 23: 547, 2000b.
- Choi, Hall, Henry, Zhang, Hein, French & Stenger, Archives of Virology 146: 619, 2001.
- Choi, Horken, Stenger,& French, Journal of General Virology 83: 443, 2002.
- Christian & Willis, Plant Disease 77: 239, 1993.
- Edwards & McMullen, North Dakota State University Farm Research Agriculture Experimental Station. Bulletin 44, 7 pp., 1987.
- Gao & Nassuth, Canadian Journal of Botany 70: 19, 1992.
- Gao & Nassuth, Journal of Phytopathology 142: 79, 1994.
- Hall, French, Hein, Morris & Stenger, Virology 282: 230, 2001.
- Harvey, Martin, & Seifers, Journal of Agricultural Entomology 11: 271, 1994.
- Hill, Martinson & Russel, Crop Science 14: 232, 1974.
- Jezewska, Phytopathologia-Polonica, 20: 77, 2000.
- Kahn & Dickerson, Phytopathology 47: 526, 1957.
- Langenberg, Journal of Ultrastructure and Molecular Structure Research 94: 161, 1986.
- Langenberg, Journal of General Virology 70: 969, 1989.
- Langenberg, Journal of Structural Biology 110: 188, 1993.
- Mahmood, Hein & French, Plant Disease 81: 250, 1997.
- McKinney, USDA Circular Number 442. 23 pp. (Washington, D. C.), 1937.
- McKinney, Plant Disease Reporter 40: 898, 1956.
- McKinney, Silber & Greely, Phytopathology 55: 1043, 1965.
- McMullen & Gardner, Journal of Ultrastructure Research 72: 65, 1980.
- McMullen, Jones, Simcox & Louie, Molecular Plant-Microbe Interactions 7: 708, 1994.
- Montana, Hunger & Sherwood, Plant Disease 80: 1239, 1996.
- Orlob, Phytopathologische Zeitschrift 55: 218, 1966.
- Paliwal, Archives of Virology 63: 123, 1980.
- Paliwal & Slykhuis, Virology 32: 344, 1967.
- Rabenstein, Seifers, Schubert, French & Stenger, Journal of General Virology 83: 897, 2002.
- Sánchez-Sánchez, Henry, Cárdenas-Soriano & Alviso-Villasana, Plant Disease 85: 13, 2001.
- Scheets, Virology 242: 28, 1998.
- Seifers, Martin, Harvey & Gill, Plant Disease 79: 1104, 1995.
- Seifers, Harvey, Kofoid & Stegmeier, Plant Disease 80: 179, 1996.
- Shepard & Carroll, Journal of Ultrastructure Research 21: 145, 1967.
- Sherwood, Journal of Phytopathology 188: 68, 1987.
- Sherwood, Hunger, Keyser & Myers, Food and Agricultural Immunology 2: 155, 1990.
- Sill & Agusioba, Plant Disease Reporter 39: 633, 1953.
- Sill & Conin, Transactions of the Kansas Academy of Science 56: 411, 1953.
- Sivamani, Brey, Dyer & Talbert, Molecular Breeding 6: 469, 2000.
- Sivamani, Brey, Talbert, Young, Dyer, Kaniewski & Qu, Transgenic Research 11: 31, 2002.
- Slykhuis, Phytopathology 45: 116, 1955.
- Slykhuis, Review of Applied Mycology 46: 401, 1967.
- Slykhuis & Bell, Canadian Journal of Botany 44: 1191, 1965.
- Staples & Allington, Research Bulletin of the Nebraska Agriculture Experimental Station 178, 40 pp., 1956.
- Stenger, Hall, Choi & French, Phytopathology 88: 782, 1998.
- Uyemoto & Ferguson, Plant Disease 64: 460, 1980.