Details of DPV and References
DPV NO: 40 October 1970
Family: Potyviridae
Genus: Potyvirus
Species: Bean yellow mosaic virus | Acronym: BYMV
Bean yellow mosaic virus
L. Bos Institute of Phytopathological Research, Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Pierce (1934).
- Selected synonyms
- Bean virus 2 (Rev. appl. Mycol. 13: 488)
- Phaseolus virus 2 (Rev. appl. Mycol. 17: 52)
- Gladiolus mosaic virus (Rev. appl. Mycol. 23: 488)
- Pea mosaic virus (Rev. appl. Mycol. 5: 337)
- Phaseolus virus 2 (Rev. appl. Mycol. 17: 52)
- A virus with flexuous particles about 750 nm long, transmitted by many aphid species in the non-persistent manner and readily by sap inoculation. It causes diseases in various cultivated and wild legumes and infects a number of non-legumes especially Liliiflorae. Most gladiolus plants are infected. It is probably common in all countries.
Main Diseases
Causes mosaic diseases in many legumes (Fig. 1 and Fig. 4); apical necrosis in French bean; ‘streak’ in pea (in part); rosette in Ornithopus sativus; white streak or latent infection in gladiolus; and unnamed diseases in fenugreek (Trigonella foenum-graecum), vetch (Vicia sativa), freesia, opium poppy (Papaver somniferum) and black locust (Robinia pseuda-acacia).
Geographical Distribution
Worldwide, in most countries where legumes have been investigated for viruses.
Host Range and Symptomatology
Formerly thought to be restricted to leguminous plants but now known to infect several non-legumes (Bos, 1970: 17 out of 20 non-legume species tested). Transmitted readily by sap inoculation or aphid vectors.
- Diagnostic species: An aid to diagnosis in most hosts is the presence
of abundant inclusion bodies of typical morphology and localization (see Relations
with Cells and Tissues).
- Phaseolus vulgaris (French bean). Inoculated primary leaves usually
epinastic with chlorotic or necrotic local lesions often extending into the
veins. Systemic yellowish mosaic (Fig. 1), leaf curling or malformation, and
often stunting. Pods sometimes mottled or malformed. Top necrosis may occur.
Symptoms greatly depend on host variety and virus strain.
Pisum sativum (pea). Transient vein chlorosis followed by mosaic, which is usually mild but, with the strains more typical of ‘pea mosaic’, may be bright yellow (Fig. 4). Sometimes necrosis occurs in stems and veins (‘streak’). ‘Perfection’ type peas are usually immune.
- Vicia faba (broad bean). Very susceptible. Transient vein chlorosis followed by obvious green or yellow mosaic. Usually no leaf distortion.
- Chenopodium amaranticolor. Inoculated leaves usually show chlorotic local lesions (Fig. 2) often followed by irregular systemic vein yellowing and leaf malformation (Fig. 3).
- Chenopodium quinoa. Chlorotic local lesions; not systemic.
- Gomphrena globosa. Local infection, often with necrotic local lesions.
- Nicotiana tabacum, Petunia hybrida, Spinacia oleracea, and Tetragonia expansa sometimes react with chlorotic local lesions; spinach may become systemically infected.
- Propagation species
- Pisum sativum, Vicia faba, Phaseolus vulgaris,
or, because of lower host protein content, Nicotiana clevelandii (systemic infection) are good sources for virus purification, and also for maintaining virus cultures.- Assay species
- Chenopodium amaranticolor
(Swenson, Sohi & Welton, 1964) and C. quinoa; some strains produce necrotic local lesions in Crotalaria spectabilis, Phaseolus aureus and some ‘Kenland’ red clover clones.
Strains
Various local strains differing in the symptoms they produce in certain varieties have been recognized, their distinction being especially important in breeding resistant varieties of crop plants, e.g.
Bean top necrosis strain of Van der Want (1954). Produces top necrosis in many bean varieties, and mosaic in various other varieties tolerant to normal strains.
Other variants more especially adapted to particular hosts are:
Pea mosaic strains. Pea mosaic virus (Doolittle & Jones, 1925) was long considered a separate virus, differing from bean yellow mosaic virus by the bright yellow symptoms it causes in pea and broad bean, and by not infecting most varieties of Phaseolus vulgaris; certain bean varieties, however, are susceptible (Schroeder & Provvidenti, 1966; Cousin, 1969). Resistance in pea to isolates of both ‘viruses’ is conditioned by the same gene (Barton et al., 1964; Cousin, 1965).
Cowpea strain of Anderson (1955) and Brierley & Smith (1962). Isolated from and produces systemic symptoms in Vigna sinensis, which is not infected by common strains of the virus. The cowpea strains give severe systemic yellow spotting in C. amaranticolor.
Red clover necrosis strain of Zaumeyer & Goth (1963). Produces necrosis in red clover and pea, local and systemic necrosis in C. amaranticolor, death of Trifolium incarnatum, T. subterraneum, and Vicia faba minor.
Transmission by Vectors
More than 20 aphid spp. can transmit the virus in a non-persistent manner (Kennedy, Day & Eastop, 1962), especially Acyrthosiphon pisum, Macrosiphum euphorbiae, Myzus persicae and Aphis fabae, although A. pisum biotypes may differ greatly in transmission efficiency (Sohi & Swenson, 1964). Aphid transmissibility of certain isolates may be lost, but this does not seem to be caused by repeated sap transmission (Swenson et al., 1964; Kamm, 1969).
Transmission through Seed
Not common. Transmitted to a small percentage in pea, Vicia faba, and white sweet clover, more in yellow and white lupin (3-6%, Zschau, 1961; 6.2% Corbett, 1958). Highest rates of transmission occur in plants infected when young.
Transmission by Dodder
No transmission by 6 Cuscuta spp. (Schmelzer, 1956).
Serology
Serological tests are difficult because it is hard to obtain sufficiently pure and concentrated virus suspensions. After intramuscular injection of the virus into rabbits titres of about 1/2048 have been obtained. These could be further increased by chemically concentrating the globulin fraction of the sera giving titres of up to 1/256,000 (Bercks, 1960, 1961), but this is exceptional. Micro-precipitin and precipitin tube tests are mostly used because intact virus particles do not diffuse in agar gel.
Relationships
Strains of bean yellow mosaic virus differ serologically to various extents, but there is no sharp distinction between closely and distantly related strains (Bercks, 1960). Pea mosaic virus is now considered a strain, even though it may differ considerably in particle length (Taylor & Smith, 1968; Bos, 1970). Bean common mosaic virus is serologically related to bean yellow mosaic virus (Beemster & Van der Want, 1951; Bercks, 1960), but differs in rarely infecting plants other than French bean and in being commonly seed-transmitted in this host. Other viruses closely related to bean yellow mosaic virus include bean western mosaic, passion fruit woodiness, cowpea aphid-borne mosaic, clover yellow vein, lupin mottle, peanut mottle, pea leafroll mosaic, pea necrosis and wisteria vein mosaic.
Bean yellow mosaic virus is distantly serologically related to potato Y and beet mosaic viruses (Bercks, 1961) and to many other members of the potato virus Y group, all of which have particles of related size and morphology. They may have more features in common than hitherto supposed; for example, several ‘non-legume’ viruses cause natural infection of legumes, and bean yellow mosaic virus infects several non-legumes. Within the potato virus Y group, there seems to exist a cluster of viruses closely related to bean yellow mosaic virus, in the same way as there is a cluster of viruses related to tobacco etch virus (Bos, 1970).
Partial, sometimes complete, cross protection in plants has been found between bean yellow mosaic and bean common mosaic viruses (Grogan & Walker, 1948; Quantz, 1961) and partial protection occurs between each virus and soybean mosaic virus (Quantz, 1961).
Stability in Sap
Depends greatly on virus source, test plant and conditions. The thermal inactivation point (10 min) is usually between 55 and 60°C but may range between 50 and 70°C, the dilution end point is usually 10-3-10-4, and longevity in vitro normally 1-2, but sometimes over 7 days.
Purification
This virus is difficult to purify because (1) particles aggregate and agglutinate with chloroplasts, so that virus is lost when sap is centrifuged or when preparations are centrifuged in sucrose density gradients, (2) particles are labile, and are broken and lost when treated with n-butanol or butanol + chloroform, and (3) preparations are contaminated with normal plant protein especially with that of legumes. Various methods have been used to remove chloroplasts and other cell debris; the best two are:
1. The ether/carbon tetrachloride method of Wetter (1960), in which either plant sap is emulsified with diethyl ether and carbon tetrachloride or plant tissue is homogenized directly into a mixture of buffer, ether and carbon tetrachloride.
2. The chloroform method of Van Regenmortel (1964), in which the tissue, after being homogenized with buffer, is emulsified in 1/5 volume of chloroform.
In both methods the emulsion is separated by low-speed centrifugation and the virus purified from the water phase by two cycles of differential centrifugation. Contaminating plant proteins can be removed by centrifuging in sucrose density gradients (Wetter, 1960), or by zone electrophoresis (Van Regenmortel, 1964).
Properties of Particles
No data.
Particle Structure
Particles are elongated and flexuous (Fig. 8), c. 750 nm long and c. 15 nm wide (Brandes & Quantz, 1955; Brandes, 1964), helically constructed with a pitch of 3.4 nm (Varma et al., 1968). Particles of greater lengths have also been reported. Bos (1970), using tobacco mosaic virus as an internal size standard, found particles 770 nm long, and Taylor & Smith (1968) and Bos (1970) found that particles from Chenopodium amaranticolor were c. 50-60 nm longer than those from legumes.
Particle Composition
No data.
Relations with Cells and Tissues
Many granular and/or crystalline cytoplasmic inclusions and often intranucleolar crystalline inclusions or nucleolar enlargements occur in epidermal tissues (Fig. 5, Fig. 6, Fig. 7) (Bos, 1969). The inclusions produced by different isolates of the virus may differ (Mueller & Koenig, 1965) but all resemble those produced by other viruses of the potato virus Y group (Bos, 1969, 1970; Edwardson, Purcifull & Christie, 1968). In ultrathin sections the cytoplasmic inclusions contain small aggregates of virus particles, electron-dense bands, pinwheel structures and crystals. The inclusions are easily stained with 0.5% trypan blue in 0.9% NaCl (McWhorter, 1941), 1% phloxine in water, or 1% phloxine alone or together with 0.2% methylene blue in a mixture of ethylene glycol monomethyl ether, ethanol and distilled water (2:1:1 v/v) (Bos, 1969).
Notes
Of several viruses closely related to bean yellow mosaic virus (see Relationships), bean common mosaic and soybean mosaic viruses are usually easily distinguished by their symptoms in French bean and soybean, respectively, and by their limited host ranges. Other closely related viruses differ slightly in host range, symptoms and inclusion bodies. They all infect several non-legumes. More distantly related members of the potato virus Y group naturally infecting legumes, such as beet mosaic, turnip mosaic and lettuce mosaic viruses, are differentiated by their infectivity towards particular non-legumes, especially their natural hosts - for details and literature, see Bos (1970).
Figures

Phaseolus vulgaris, ‘Processor’, systemically infected.

Lesions in inoculated leaf of Chenopodium amaranticolor.

Systemic symptoms in C. amaranticolor.

Systemic symptoms in Pisum sativum, ‘Koroza’, produced by (left) a bean isolate and (right) a pea isolate.
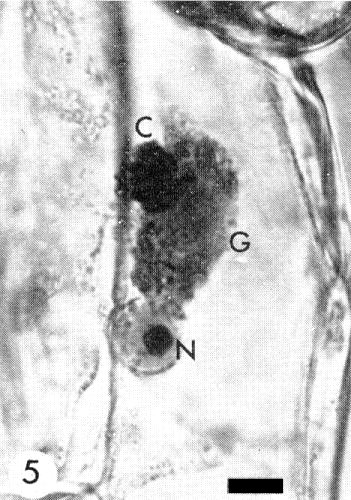
Cytoplasmic crystalline (C) and granular (G) inclusions and intranucleolar (N) inclusions induced by a pea isolate in Vicia faba. Bar represents 0.01 mm.
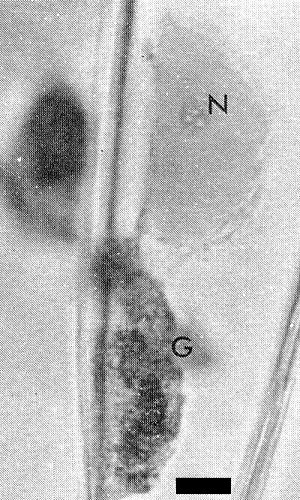
Cytoplasmic granular (G) inclusions and intranucleolar (N) inclusions induced by a pea isolate in Vicia faba. Bar represents 0.01 mm.
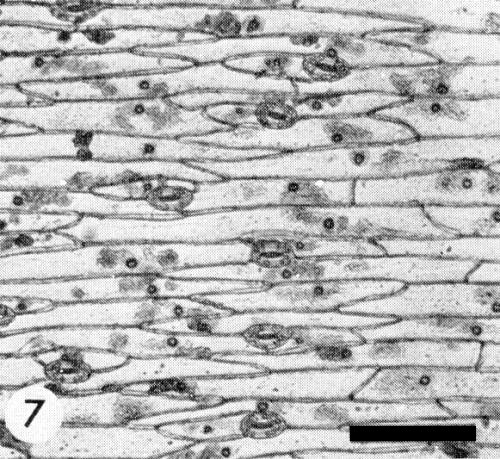
Vicia faba petiole epidermis with inclusion bodies. Bar represents 0.1 mm.
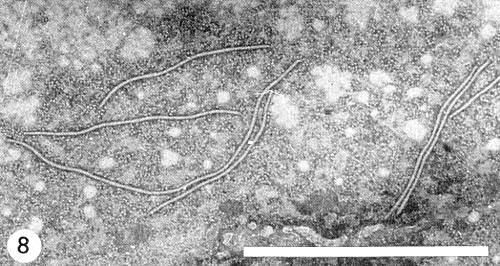
Electron micrograph of virus particles from pea, cut-leaf preparations stained with phosphotungstate. Bar represents 1 µm. Note end-to-end aggregation and subsequent production of particles with abnormal lengths (the second particle from top left measures about 900 nm).
References list for DPV: Bean yellow mosaic virus (40)
- Anderson, Pl. Dis. Reptr 39: 349, 1955.
- Barton, Schroeder, Provvidenti & Mishanec, Pl. Dis. Reptr 48: 353, 1964.
- Beemster & Van der Want, Antonie van Leeuwenhoek 17: 15, 1951.
- Bercks, Phytopath. Z. 39: 120, 1960.
- Bercks, Phytopath. Z. 40: 357, 1961.
- Bos, Neth. J. Pl. Path. 75: 137, 1969.
- Bos, Neth. J. Pl. Path. 76: 8, 1970.
- Brandes, Mitt. biol. Bund Anst. Ld-u. Forstw. (Berlin-Dahlem) 110, 130 pp., 1964.
- Brandes & Quantz, Naturwissenschaften 42: 588, 1955.
- Brierley & Smith, Pl. Dis. Reptr 46: 335, 1962.
- Corbett, Phytopathology 48: 86, 1958.
- Cousin, Annls Amél. Pl. 15: 23, 1965.
- Cousin, Annls Phytopath. 1: 210, 1969.
- Doolittle & Jones, Phytopathology 15: 763, 1925.
- Edwardson, Purcifull & Christie, Virology 34: 250, 1968.
- Grogan & Walker, Phytopathology 38: 489, 1948.
- Kamm, Ann. ent. Soc. Am. 62: 47, 1969.
- Kennedy, Day & Eastop, A conspectus of aphids as vectors of plant viruses, London, Commonwealth Institute of Entomology, 1962.
- McWhorter, Stain Technol. 16: 143, 1941.
- Mueller & Koenig, Phytopathology 55: 242, 1965.
- Pierce, Phytopathology 24: 87, 1934.
- Quantz, Phytopath. Z. 43: 79, 1961.
- Schmelzer, Phytopath. Z. 28: 1, 1956.
- Schroeder & Provvidenti, Pl. Dis. Reptr 50: 337, 1966.
- Sohi & Swenson, Entomologia exp. appl. 7: 9, 1964.
- Swenson, Sohi & Welton, Ann. ent. Soc. Am. 57: 378, 1964.
- Taylor & Smith, Aust. J. biol. Sci. 21: 429, 1968.
- Van der Want, Meded. Inst. plziektenk. Onderz. 85: 1,1954.
- Van Regenmortel, Virology 23: 495, 1964.
- Varma, Gibbs, Woods & Finch, J. gen. Virol. 2: 107, 1968.
- Wetter, Arch. Mikrobiol. 37: 278, 1960.
- Zaumeyer & Goth, Pl. Dis. Reptr 47: 10, 1963.
- Zschau., NachrBl. dt. PflSchutzdienst, Berl. 15: 1, 1961.