Details of DPV and References
DPV NO: 400 July 2003
Family: Bromoviridae
Genus: Cucumovirus
Species: Cucumber mosaic virus | Acronym: CMV
This is a revised version of DPV 213
Cucumber mosaic virus
Peter Palukaitis Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, UK
Fernando García-Arenal Depto. De Biotecnologia, E.T.S.I. Agrónomos, Universidad Politécnica de Madrid, 28040 Madrid, Spain
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Doolittle (1916) and Jagger (1916).
- Selected synonyms
- Coleus mosaic virus (Creager, 1945)
- Lily ringspot virus (Brierley & Travis, 1958)
- Southern celery mosaic virus (Doolittle, 1916)
- Soybean stunt virus (Hanada & Tochihara, 1982)
- At least 43 others (Kaper & Waterworth, 1981)
Brief description
A virus with a three-component genome of ss RNA of messenger sense. RNA is packaged in three different
icosahedral particles, 30.5 nm (maximum) in diameter, which sediment at the same rate. In nature, it is
transmitted by aphids in a non-persistent manner, and through the seed in some plant hosts. Experimentally,
it is easily transmitted by mechanical inoculation of plant sap. It is distributed world-wide, causes
economically important diseases in a large variety of crop plants, and has the widest host range of any
known plant virus.
Main Diseases
Causes mosaic in cucumber, melon and other cucurbits; blight in spinach; mosaic, fern leaf and systemic necrosis in tomato; mosaic and ringspot in pepper; mosaic and stunting in clover, lupins and lucerne; stunting in soybean; mosaic, infectious chlorosis and heart rot in banana; and mosaic and dwarfing in many other species of dicotyledonous and monocotyledonous plants (Kaper & Waterworth, 1981; Palukaitis et al., 1992; Brunt et al., 1997). The presence of satellite RNA can affect the level of CMV replication and its pathogenicity. Notably, systemic necrosis of tomato, reported in France (Marrou et al., 1973), Italy (Gallitelli et al., 1988), Spain (Jordá et al., 1992) and Japan (Yoshida et al., 1984) is due to the presence of satellite RNAs that induce necrosis in this host plant.
Geographical Distribution
World-wide, both in tropical and temperate climates.
Host Range and Symptomatology
CMV has the widest host range for any plant virus, including more than 1200 species in over 100 families of dicotyledonous and monocotyledonous angiosperms (Douine et al., 1979; Edwardson & Christie, 1991). Induces a variety of symptoms depending on the host plant species and cultivar, and on the virus strain. Some strains induce severe chlorosis.
- Diagnostic species
Beta vulgaris. Large chlorotic local lesions; not systemic. (Fig. 1)
Chenopodium amaranticolor and C. quinoa. Chlorotic or necrotic local lesions. Rarely systemic. (Fig. 2)
Cucumis sativus (cucumber). Systemic mosaic and stunting varying in severity with the virus strain. (Fig. 3)
Cucurbita spp. (zucchini squash, marrow or courgette). Systemic mosaic and stunting of varying severity. (Fig. 4)
Lycopersicon esculentum (tomato). Mosaic and stunting with filiform leaves to different extents, often extreme (fern leaf). (Fig. 5)
Nicotiana tabacum. Mild to severe mosaic and stunting, depending on the virus strain. (Fig. 6) Some strains induce severe yellow chlorosis. (Fig. 7) In inoculated leaves, most isolates of Subgroup I (A or B) do not induce symptoms, but most isolates in Subgroup II induce etched rings (Fig. 8), as well as in systemically infected leaves in some instances (due possibly to environmental effects). Isolates of Subgroup I that infect Vigna spp. systemically may induce chlorotic lesions in inoculated leaves of tobacco.
Phaseolus aureus and P. vulgaris cv. Pinto. Small purple necrotic lesions in inoculated leaves.
Vigna unguiculata (cowpea). Large (isolates of Subgroup I A or B) (Fig. 9) or small (isolates of Subgroup II) (Fig. 10) brown lesions in inoculated leaves. Only some Subgroup I isolates are systemic causing mild mosaic and these induce chlorotic lesions in inoculated leaves.
Nicotiana benthamiana. Severe stunting, leaf curling and leaf deformation, some strains also produce necrosis along veins.
- Propagation species
Nicotiana tabacum or N. glutinosa are convenient for maintaining cultures. Cucurbita spp. and N. tabacum are suitable as virus source plants for purifying the virus, but N. clevelandii appears the best source for most strains.
- Assay species
Vigna unguiculata, Chenopodium amaranticolor and C. quinoa are suitable for local lesion assays. For transmission studies, cucumber is suitable for Aphis gossypii, and N. clevelandii for Myzus persicae.
- Abdalla, Desjardins & Dodds, Plant Disease 75: 1019, 1991.
- Ahlquist, Dasgupta & Kaesberg, Cell 23: 183, 1981.
- Alonso-Prados, Fraile & García-Arenal, Journal of Plant Pathology 79: 131, 1997.
- Anonymous, Journal of Plant Pathology 80: 133, 1998.
- Badami, Annals of Applied Biology 46: 554, 1958.
- Bao, Flasinski & Roossinck, in Abstracts of the XIth International Congress of Virology, 9-13 August 1999, Sydney, Australia, 304, 1999 (Abstract).
- Bernal & García-Arenal, RNA 3: 1052, 1997.
- Blackman, Boevink, Santa Cruz, Palukaitis & Oparka, Plant Cell 10: 525, 1998.
- Blanchard, Boyce & Anderson, Virology 217: 598, 1996.
- Bouhida & Lockhart, Phytopathology 80: 981, 1990.
- Bouwen, Witt & Dijkstra, Netherlands Journal of Plant Pathology 84: 175, 1978.
- Brierley & Travis, Plant Disease Reporter 42: 1034, 1958.
- Brigneti, Voinnet, Li, Ji, Ding & Baulcombe, EMBO Journal 17: 6739, 1998.
- Brunt, Crabtree, Dallwitz, Gibbs, Watson & Zurcher, Plant Viruses Online: Descriptions and lists from the VIDE database. Version 16th January 1997.
- Bwye, Proudlove, Berlandier & Jones, Australian Journal of Experimental Agriculture 37: 1, 1997.
- Canto & Palukaitis, Virology 265: 74, 1999a.
- Canto & Palukaitis, Molecular Plant-Microbe Interactions 12: 985, 1999b.
- Canto, Prior, Hellwald, Oparka & Palukaitis, Virology 237: 237, 1997.
- Chen & Francki, Journal of General Virology 71: 939, 1990.
- Chen, Randles & Francki, Journal of General Virology 76: 971, 1995.
- Choi, Choi, Park & Ryu, Journal of Virological Methods 83: 67, 1999.
- Christie & Edwardson, Florida Agricultural Experiment Station Monograph Series No.9, 89, 1977.
- Creager, Phytopathology 35: 223, 1945.
- Crescenzi, Barbarossa, Gallitelli & Martelli, Plant Disease 77: 28, 1993a.
- Crescenzi, Barbarossa, Cillo, Di Franco, Vovlas & Gallitelli, Archives of Virology 131: 321, 1993b.
- Davies & Symons, Virology 165: 216, 1988.
- Devergne & Cardin, Annales de Phytopathologie 5: 409, 1973.
- Ding, Anderson, Haase & Symons, Virology 198: 593, 1994.
- Ding, Li & Symons, EMBO Journal 14: 5762, 1995a.
- Ding, Li, Nguyen, Palukaitis & Lucas, Virology 207: 345, 1995b.
- Ding, Shi, Li & Symons, Proceedings of the National Academy of Sciences of the United States of America 93: 7470, 1996.
- Doolittle, Phytopathology 6: 145, 1916.
- Douine, Quiot, Marchoux & Archange, Annales de Phytopathologie 11: 439, 1979.
- Edwards & Gonsalves, Phytopathology 73: 1117, 1983.
- Edwardson & Christie, CRC Handbook of viruses infecting legumes, Boca Raton, CRC Press, 293, 1991.
- Ehara & Mink, Tohoku Journal of Agricultural Research 34: 31, 1984.
- Finch, Klug & Van Regenmortel, Journal of Molecular Biology 24: 303, 1967.
- Fraile, Alonso-Prados, Aranda, Bernal, Malpica & García-Arenal, Journal of Virology 71: 934, 1997.
- Francki, Virology 24: 193, 1964.
- Francki, Virology 34: 694, 1968.
- Francki & Habili, Virology 48: 309, 1972.
- Francki & Hatta, Acta Horiculturae 110: 167, 1980.
- Francki, Mossop & Hatta, CMI/AAB Description of Plant Viruses 213, 1979.
- Francki, Randles, Chambers & Wilson, Virology 28: 729, 1966.
- Fuchs, Tricoli, Carney, Schesser, McFerson & Gonsalves, Plant Disease 82: 1350, 1998.
- Gallitelli, Di Franco, Vovlas & Kaper, Informatore Fitopatologico 12: 57, 1988.
- Gal-On, Kaplan, Roossinck & Palukaitis, Virology 205: 280, 1994.
- Gal-On, Wolf, Wang, Faure, Pilowsky & Zelcer, Phytopathology 88: 1101, 1998.
- Gera, Loebenstein & Raccah, Phytopathology 69: 396, 1979.
- Gerola, Bassi & Belli, Caryologia 18: 567, 1965.
- Gillaspie, Hajimorad & Ghabrial, Plant Disease 82: 419, 1998.
- Gonda & Symons, Virology 88: 361, 1978.
- Gonsalves, Provvidenti & Edwards, Phytopathology 72: 1533, 1982.
- Gordon & Symons, Nucleic Acids Research 11: 947, 1983.
- Grafton-Cardwell, Perring, Smith, Valencia & Farrar, Plant Disease 80: 1092, 1996.
- Habili & Francki, Virology 61: 443, 1974a.
- Habili & Francki, Virology 57: 392, 1974b.
- Hampton & Francki, Phytopathology 82: 127, 1992.
- Hanada & Tochihara, Phytopathology 72: 761, 1982.
- Hatta & Francki, Virology 93: 265, 1979.
- Hatta & Francki, Journal of General Virology 53: 343, 1981.
- Havey, Cucurbit Genetics Cooperative Report.20: 18, 1997.
- Hayes & Buck, Cell 63: 363, 1990.
- Hidaka & Tomaru, Bulletin of the Hatano Tobacco Experiment Station 46: 125, 1960.
- Hollings & Stone, CMI/AAB Descriptions of Plant Viruses 79, 1971.
- Honda & Matsui, Phytopathology 58: 1230, 1968.
- Honda & Matsui, Phytopathology 64: 534, 1974.
- Hsu, Wu, Lin, Chen, Lee & Tsai, Archives of Virology 140: 1841, 1995.
- Hu & Ghabrial, Journal of Virological Methods 55: 367, 1995.
- Jagger, Phytopathology 6: 148, 1916.
- Jaspars, Gill & Symons, Virology 144: 410, 1985.
- Ji & Ding, Molecular Plant-Microbe Interactions 14: 715, 2001.
- Jones & Coutts, Annals of Applied Biology 129: 491, 1997.
- Jones & McLean, Annals of Applied Biology 114: 609, 1989.
- Jordá, Alfaro, Aranda, Moriones & García-Arenal, Plant Disease 76: 363, 1992.
- Kaper, Virology 37: 134, 1969.
- Kaper, The Chemical Basis of Virus Structure, Dissociation and Reassembly, New York Elsevier/North Holland, 485 pp, 1975.
- Kaper & Geelen, Journal of Molecular Biology 56: 227, 1971.
- Kaper & Re, Virology 60: 308, 1974.
- Kaper & Waterworth, in Handbook of Plant Virus Infection and Comparative Diagnosis, p. 257, ed. Kurstak, New York, Elsevier/North-Holland, 1981.
- Kaper, Tousignant & Lot, Biochemical and Biophysical Research Communications 72: 1273, 1976.
- Kaplan & Palukaitis, Virology 251: 729, 1998.
- Kaplan, Gal-On & Palukaitis, Virology 230: 343, 1997.
- Kaplan, Zhang & Palukaitis, Virology 246: 221, 1998
- Kaplan, Shintaku, Li, Zhang, Marsh & Palukaitis, Virology 209: 188, 1995.
- Karasawa, Ito, Okada, Hase & Ehara, Annales of the Phytopathological Society of Japan 63: 289, 1997.
- Karasawa, Okada, Akashi, Chida, Hase, Nakazawa-Nasu, Ito & Ehara, Phytopathology 89: 1186, 1999.
- Kataoka, Masuta & Takanami, Annales of the Phytopathological Society of Japan 56: 495, 1990a.
- Kataoka, Masuta & Takanami, Annales of the Phytopathological Society of Japan 56: 501, 1990b.
- Kim & Palukaitis, EMBO Journal 16: 4060, 1997.
- Kobori, Miyagawa, Nishioka, Ohki & Osaki, Journal of General Plant Pathology 68: 81, 2002.
- Kohl & Hall, Journal of General Virology 25: 257, 1974.
- Lakshman & Gonsalves, Phytopathology 75: 758, 1985.
- Li, Lucy, Guo, Li, Ji, Wong & Ding, EMBO Journal 18: 2683, 1999.
- Li, Ryu & Palukaitis, Molecular Plant-Microbe Interactions 14: 378, 2001.
- Liu, He, Park, Josefsson & Perry, Journal of Virology 76: 9756, 2002.
- Lot & Kaper, Virology 74: 209, 1976a.
- Lot & Kaper, Virology 74: 223, 1976b.
- Lot, Marrou, Quiot & Esvan, Annales de Phytopathologie 4: 25, 1972.
- Lot, Marchoux, Marrou, Kaper, West, Van Vloten-Doting & Hull, Journal of General Virology 22: 81, 1974.
- Luis-Arteaga, Rodríguez-Cerezo, Maestro & García-Arenal, Plant Disease 72: 265, 1988.
- Luis-Arteaga, Alvarez, Alonso-Prados, Bernal, García-Arenal, Laviña, Batlle & Moriones, Plant Disease 82: 979, 1998.
- Lyons & Allen, Journal of Ultrastructure Research 27: 198, 1969.
- Marchoux, Acta Horticulturae 227: 57, 1990.
- Marchoux, Doine, Marrou & Devergne, Annales de Phytopathologie 4: 363, 1972.
- Marchoux, Marrou, Devergne, Quiot, Douine & Lot, Mededelingen Faculteit Landbouwwetenschappen Rijksuniversiteit Gent 40: 59, 1975.
- Marrou, Duteil, Lot & Clerjeau, Pépiniérstes Horticulteurs Maraîchers 137: 37, 1973.
- Martelli & Russo, in Plant Viruses. Vol.1. Polyhedral Virions with Tripartite Genomes, ed. Francki, New York: Plenum Press, p.163, 1985.
- Masuta, Ueda, Suzuki & Uyeda, Proceedings of the National Academy of Sciences of the United States of America 95: 10487, 1998.
- Mayers, Palukaitis & Carr, Journal of General Virology 81: 219, 2000.
- McGarvey, Tousignant, Geletka, Cellini & Kaper, Journal of General Virology 76: 2257, 1995.
- Mink, CMI/AAB Descriptions of Plant Viruses 92, 1972.
- Mossop & Francki, Virology 81: 177, 1977.
- Mossop & Francki, Virology 95: 395, 1979.
- Mossop, Francki & Grivell, Virology 74: 544, 1976.
- Nagano, Mise, Furusawa & Okuno, Journal of Virology 75: 8045, 2001.
- Ng & Perry, Plant Pathology 48: 388, 1999.
- Ng, Liu & Perry, Virology 276: 395, 2000.
- Nitta, Masuta, Kuwata & Takanami, Annales of the Phytopathological Society of Japan 54: 516, 1988.
- Owen & Palukaitis, Virology 166: 495, 1988.
- Owen, Shintaku, Aeschleman, Ben Tahar & Palukaitis, Journal of General Virology 71: 2243, 1990.
- Palukaitis & García-Arenal, in Current Topics in Microbiology and Immunology 239: 37, 1999.
- Palukaitis, Roossinck, Dietzgen & Francki, Advances in Virus Research 41: 281, 1992.
- Peden & Symons, Virology 53: 487, 1973.
- Perry & Francki, Journal of General Virology 73: 2105, 1992.
- Perry, Habili & Dietzgen, Plant Pathology 42: 806, 1993.
- Perry, Zhang & Palukaitis, Virology 242: 204, 1998.
- Perry, Zhang, Shintaku & Palukaitis, Virology 205: 591, 1994.
- Piazzolla, Díaz-Ruíz & Kaper, Journal of General Virology 45: 361, 1979.
- Pitrat & Lecoq, Phytopathology 70: 958, 1980.
- Price, Phytopathology 24: 743, 1934.
- Provvidenti, Plant Disease Reporter 60: 289, 1976.
- Provvidenti, Robinson & Shail, Hortscience 15: 528, 1980.
- Quiot, Labonne & Quiot-Douine, in Plant Virus Epidemiology, p.1777, eds Plumb & Thresh, Oxford, Blackwell, 1983.
- Rao & Francki, Virology 114: 573, 1981.
- Rao & Francki, Journal of General Virology 61: 197, 1982.
- Rezaian, Williams, Gordon, Gould & Symons, European Journal of Biochemistry 143: 277, 1984.
- Rezaian, Williams & Symons, European Journal of Biochemistry 150: 331, 1985.
- Rist & Lorbeer, Phytopathology 81: 367, 1991.
- Rizzo & Palukaitis, Journal of General Virology 69: 1777, 1988.
- Rizzo & Palukaitis, Journal of General Virology 70: 1, 1989.
- Rodríguez-Alvarado & Roossinck, Virology 236: 155, 1997.
- Roossinck, Journal of Virology 76: 3382, 2002.
- Roossinck & Palukaitis, Molecular Plant-Microbe Interactions 3: 188, 1990.
- Roossinck, Kaplan & Palukaitis, Journal of Virology 71: 608, 1997.
- Roossinck, Zhang & Hellwald, Journal of Virology 73: 6752, 1999.
- Ryabov, Roberts, Palukaitis & Taliansky, Virology 260: 98, 1999.
- Ryu, Kim & Palukaitis, Molecular Plant-Microbe Interactions 11: 351, 1998.
- Sackey & Francki, Physiological and Molecular Plant Pathology 36: 409, 1990.
- Saitoh, Fujiwara, Ohki & Osaki, Annales of the Phytopathological Society of Japan 65: 248, 1999.
- Salánki, Thole, Balázs & Burgyán, Virus Research 31: 379, 1994.
- Schmelzer, Phytopathologishe Zeitschrfit 28: 1, 1957.
- Schmitz & Rao, Virology 248: 323, 1998.
- Schwinghamer & Symons, Virology 63: 252, 1975.
- Scott, Virology 34: 79, 1968.
- Shi, Palukaitis & Symons, Molecular Plant-Microbe Interactions 15: 947, 2002.
- Shintaku, Zhang & Palukaitis, Plant Cell 4: 751, 1992.
- Simons, Phytopathology 45: 217, 1955.
- Simons, Phytopathology 47: 145, 1957.
- Singh, Jones & Jones, Plant Disease 79: 713, 1995.
- Smith, Chase, Schmidt & Perry, Journal of Virology 74: 7578, 2000.
- Soards, Murphy, Palukaitis & Symons, Molecular Plant-Microbe Interactions 15: 647, 2002.
- Sugiyama, Sato, Karasawa, Hase, Takahashi & Ehara, Physiological and Molecular Plant Pathology 56: 85, 2000.
- Suzuki, Kuwata, Masuta & Takanami, Journal of General Virology 76: 1791, 1995.
- Symons, Molecular Biology Reports 2: 277, 1975.
- Symons, Nucleic Acids Research 7: 825, 1979.
- Szilassy, Salánki & Balázs, Molecular Plant-Microbe Interactions 12: 1105, 1999a.
- Szilassy, Salánki & Balázs, Virus Genes 18: 221, 1999b.
- Takahashi, Sugiyama, Sukamto, Karasawa, Hase & Ehara, Journal of General Plant Pathology 66: 335, 2000.
- Takeshita, Suzuki, Kuwata & Takanami, Archives of Virology 143: 1109, 1998.
- Takeshita, Suzuki & Takanami, Archives of Virology 146: 697, 2001.
- Tien & Wu, Advances in Virus Research 39: 321, 1991.
- Tomaru & Hidaka, Bulletin of the Hatano Tobacco Experiment Station 46: 143, 1960.
- Van Regenmortel, Virology 31: 391, 1967.
- Van Regenmortel, Hendry & Baltz, Virology 49: 647, 1972.
- Vaquero, Liao, Nähring & Fischer, Journal of General Virology 78: 2095, 1997.
- Wahyuni, Dietzgen, Hanada & Francki, Plant Pathology 41: 282, 1992.
- Waterworth & Povish, Phytopathology 65: 728, 1975.
- White, Morales & Roossinck, Virology 207: 334, 1995.
- Wickoff, Tsai, Wang, Baker & Johnson, Virology 232: 91, 1997.
- Wong, Thio, Shintaku & Palukaitis, Molecular Plant-Microbe Interactions 12: 628, 1999.
- Yang, Kim & Anderson, Phytopathology 87: 924, 1997.
- Yoshida, Goto & Tizuka, Annals of the Phytopathological Society of Japan 50: 92, 1984.
- Zhang, Hanada & Palukaitis, Journal of General Virology 75: 3185, 1994.
- Zehen & Berger, Phytopathology 80: 769, 1990.
- Ziegler, Torrance, Macintosh, Cowan & Mayo, Virology 214: 235, 1995.
Strains
Many symptom variants occur, making the virus often difficult to identify from symptoms alone. Many strains have been differentiated on the basis of symptom and host range, transmission, serology, physicochemical properties and nucleotide sequence. Most strains have a restricted geographic distribution. Among the many well-characterised strains are Price's No. 6 (Price, 1934), O (Hidaka & Tomaru, 1960) Y (Tomaru & Hidaka, 1960), Q (Francki et al., 1966), S (Van Regenmortel, 1967), D (Marchoux et al., 1972), Ix (Waterworth & Povish, 1975), M (Mossop et al., 1976), B (Provvidenti, 1976), LS (Provvidenti et al., 1980), WL (Gonsalves et al., 1982), Fny (Roossinck & Palukaitis, 1990), Tfn (Crescenzi et al., 1993b) and NT9 (Hsu et al., 1995).
Transmission by Vectors
Transmissible in a non-persistent manner by more than 80 species of aphids in 33 genera (Edwardson & Christie, 1991). Myzus persicae and Aphis gossypii are two important vectors. Transmission efficiency is affected by a number of factors, including virus strain, aphid species, source and recipient plant species, and plant species on which the aphid is maintained (Simons, 1957). Virus is acquired and transmitted by all instars within 5-10 sec of feeding. Transmission ability declines after feeding for about 2 min and is usually lost within 4 h (Simons, 1955), although this short retention time could be an artefact of experimental design (Zehen & Berger, 1990). Virus isolates are known that have lost the ability to be aphid transmissible (Badami, 1958; Mossop & Francki, 1977; Ng & Perry, 1999), but the loss of transmissibility upon repeated mechanical passage seems to be rare (Ng & Perry, 1999). Differences in transmissibility of various isolates by different aphid species are determined by the virus coat protein (Gera et al., 1979; Chen & Francki, 1990) and amino acid determinants for transmission have been mapped (Perry et al., 1994, 1998; Liu et al., 2002).
Transmission through Seed
Seed transmission has been reported in more than twenty plant species, with varying efficiencies from a fraction of 1% up to 50% (Palukaitis et al., 1992). Virus may be present in the embryo, endosperm and seminal integuments, as well as in pollen (Yang et al., 1997). Seed transmission in weeds is of epidemiological significance (Quiot et al., 1983; Rist & Lorbeer, 1991).
Transmission by Dodder
Transmitted by at least ten species of Cuscuta, in which the virus multiplies (Schmelzer, 1957).
Serology
The virus is poorly immunogenic, but this can be enhanced by fixation with formaldehyde (Francki & Habili, 1972) or glutaraldehyde (Francki & Hatta, 1980). Because the virus precipitates on exposure to physiological salt solutions and mild heating, gel immunodiffusion tests are done usually in buffers of low molarity or water (Francki et al., 1966; Scott, 1968). Polyclonal antibodies show different results depending on the type of ELISA used: plate-trapped antigen ELISA shows less strain specificity than double antibody sandwich ELISA (Anonymous, 1998), but gives similar overall results to gel immunodiffusion tests (Wahyuni et al., 1992). Antisera can vary considerably in their ability to detect virus in ELISA and in immunoblots. Several panels of monoclonal antibodies have been prepared with variation in specificity. The specificity can vary also with the technique used (type of ELISA or type of immunoblot). Single-chain variable fragment antibody to CMV has been prepared from a synthetic phage display library (Ziegler et al., 1995).
Relationships
Strains of CMV have been classified on the basis of serological typing (Devergne & Cardin, 1973), peptide mapping of the coat protein (Edwards & Gonsalves, 1983) and sequence similarity of their genomic RNAs (Gonda & Symons, 1978, Piazzolla et al., 1979, Owen & Palukaitis, 1988). These classifications divided CMV strains into either Subgroup I or Subgroup II (Owen & Palukaitis, 1988). Overall nucleotide sequence similarity among isolates is 90% to 98% within Subgroup I and about 98% within Subgroup II; similarities between isolates from different subgroups is 65% to 70%. Analysis of RNA 3 sequences of 26 isolates indicates that Subgroup I strains can be subdivided further into Subgroups IA and IB, in which Subgroup IA strains are more closely clustered than are Subgroup IB strains. Subgroups IA and II appear to be monophyletic (Roossinck et al., 1999). Similar relationships are obtained comparing sequences of RNAs 1 or 2 (Roossinck, 2002). Cross protection occurs between strains from all subgroups. CMV is related to Tomato aspermy virus (TAV, Hollings & Stone, 1971) and to Peanut stunt virus (PSV, Mink, 1972) the two other definite members of the genus Cucumovirus; Robinia mosaic virus is now considered a strain of PSV. Nucleotide sequence similarity amongst these three cucumoviruses is about 60% to 65% and they can be differentiated serologically and by host range (Kaper & Waterworth, 1981). Monoclonal antibodies have been obtained that can distinguish CMV Subgroup I and Subgroup II isolates, and/or react with most of the isolates (Wahyuni et al., 1992; Anonymous, 1998). RT-PCR has been used to detect cucumoviruses and differentiate isolates in the CMV subgroups (Anonymous, 1998; Choi et al., 1999).
Pseudo-recombinant viruses that multiply efficiently can be obtained by exchange of genomic segments between CMV isolates from both subgroups, and to a lesser extent, between CMV and the other cucumoviruses (Habili & Francki, 1974a; Marchoux et al., 1975; Rao & Francki, 1981, 1982; Sackey & Francki, 1990; Masuta et al., 1998). Pseudo-recombinants are also isolated by aphid transmission from mixed infections (Perry & Francki, 1992; Fraile et al., 1997). Nevertheless, pseudo-recombinants between Subgroups I and II have not been reported in nature, and those produced between strains in the same subgroup are infrequent (Fraile et al., 1997). A natural pseudo-recombinant between CMV and PSV has been reported (White et al., 1995).
Stability in Sap
Relatively unstable in plant sap. Thermal inactivation point (10 min) is 55-70 °C. Infectivity can be lost in a few hours or in a few (less than 10) days. Infectivity is inhibited by leaf extracts from all plants tested (Francki, 1964), and is abolished by ribonuclease treatment (Francki, 1968). Virus cultures can be preserved as freeze-dried tissues from infected plants, as purified virions in 5mM sodium borate, 0.5mM EDTA, pH 9.0, 50% glycerol at -20 °C, or as purified virus RNAs stored at -20 °C.
Purification
Most strains can be purified following the procedure originally described by Lot et al. (1972). Infected leaves are blended in an equal volume of 0.5M sodium citrate, pH 6.5, containing 0.5% thioglycolic acid. The homogenate is clarified with 1 vol chloroform, and centrifuged at 12,000 g for 10 min. Virus in the aqueous phase is precipitated by adding polyethylene glycol (mol. wt 8,000) to 10%, gently stirring for 30-45 min at 0-4 °C, and centrifuging at 7,000 g for 20 min. The pellet is resuspended in 50mM sodium citrate buffer, pH 7.0 containing 2% Triton X-100 and stirred for 30 min before centrifuging at 19,000 g for 15 min. Further purification is achieved by cycles of differential centrifugation and/or centrifuging in 5-25% sucrose density gradients. The purified virus particle preparation is resuspended in 50mM sodium citrate pH 7.0 or 50mM sodium citrate with 5mM sodium borate, 0.5 mM EDTA, pH 9.2. For the few strains that cannot be purified by this method, an alternative method uses phosphate buffer for extraction, and avoids the use of organic solvents, as described by Mossop et al. (1976). However, most strains aggregate in phosphate buffers. Virus yields depend on the virus isolate, the presence or absence of satellite RNA, and the source plant, but are usually 0.1-1 g/kg.
Properties of Particles
Particles are stabilised by RNA-protein interactions, and are readily disrupted in low concentrations of sodium dodecyl sulphate or high concentrations of neutral chloride salts (Kaper, 1975).
Sedimentation coefficient: particles sediment as a single component with S20,w (svedbergs) of 98.6-104 c (c being virus concentration in mg m1-1) (Francki et al., 1966).
Buoyant density (g.cm-3): 1:36 in CsC1, after formaldehyde fixation of particles; 1.34 in RbCl (Lot & Kaper, 1976a).
Particle weight: Estimates vary from 5.0 to 6.7 x 106 (Francki et al., 1979). Based on the known sequences of the viral RNAs, the particle Mol. Wt should be c. 5.6 x 106.
Diffusion coefficient: D20,w (x 10-7 cm2.sec-1): 1.29-1.35 for four strains using laser light scattering (Francki et al., 1979).
Isoelectric point: 4.75 (Ehara & Mink, 1984).
Partial specific volume (cm3.g -1): 0.701 (Francki et al., 1979).
Electrophoretic mobility: varies with the strain (Lot & Kaper, 1976a). Strain S is - 8 x 10-5 cm2sec-1V-1 at pH 7.0 in 0.1 M buffer. Mobility as a function of pH is given by Van Regenmortel (1967).
Extinction coefficient at 260 nm (1 mg.m1-1, 1cm light path): 5.0 (Francki et al., 1966).
A260/A280: about 1.7, corrected for light scattering (Francki et al., 1979).
Particle Structure
Particles are isometric, about 28-30 nm in diameter, as shown when negatively stained in uranyl acetate pH 4.5 (Francki & Hatta, 1980). (Fig. 11) Particles are also stable in ammonium molybdate (Bouwen et al., 1978), but disrupt if mounted unfixed in neutral phosphotungstate (Francki et al., 1966). Particles contain 180 protein subunits in pentamer-hexamer clusters with T = 3 symmetry (Finch et al., 1967). The three dimensional structure of particles has been analysed at 3.2 Å resolution by X-ray chrystallography (Smith et al., 2000). The exterior radius along the quasi-sixfold axes is 144 Å, and the density along the five-fold axes extends 3 Å further. The electron density map for each protein subunit is consistent with a beta-barrel motif. The long axis of the beta-barrel domain is oriented in a roughly radial direction. This structure is very similar to the better-characterised one of the Cowpea chlorotic mottle virus, genus Bromovirus (Smith et al., 2000). The RNA is packed tightly against the protein shell leaving a hollow core of about 110 Å along the three-fold axes. Particles are stabilised mainly by protein-RNA interactions (Kaper & Geelen, 1971) that occur between the negatively charged phosphate groups of the RNA and, probably, the positively charged Arg residues in the N-terminus of the coat protein (Wickoff et al., 1997; Smith et al., 2000). Particles can be dissociated readily into their components and reassembled into biologically active particles (Kaper, 1969). Reassembly of particles formed of either CMV or TAV coat proteins also is possible (Chen et al., 1995).
Particle Composition
Nucleic acid: Single stranded messenger sense RNA, that is about 18% of particle weight (Kaper & Re, 1974). The G:A:C:U ratios are c. 24:23:23:30 (Habili & Francki, 1974b). RNA consists of four major species designated 1, 2, 3 and 4, containing c. 3,350, 3,050, 2,020, and 1,030 nt, respectively, with variation in nucleotide numbers found primarily in the 5' nontranslated region of RNA 3 in different isolates (Roossinck et al., 1999). Only the largest three RNAs are required for infectivity; RNA 4 is the subgenomic messenger for the coat protein, derived from RNA 3 (Peden & Symons, 1973; Lot et al., 1974). RNA 1 and RNA 2 are encapsidated separately, whereas RNA 3 and RNA 4 are probably packaged together in the same particle (Habili & Francki, 1974b; Lot & Kaper, 1976b). Some particles may exist containing three molecules of RNA 4 (Kaplan & Palukaitis, 1998). Thus virus preparations contain at least three different types of particles, but with similar morphology and sedimentation properties. Virus particles also contain low levels of smaller sized RNA species, designated RNA 4A, RNA 5 and RNA 6. RNA 4A of 682 nt in strain Q is derived from RNA 2 and is the subgenomic messenger for gene 2b (Ding et al., 1994). RNA 5 of 309 nt in strain Q, is a heterogeneous mixture of the 3' noncoding regions of RNA 2 and RNA 3 (Blanchard et al., 1996). RNA 6 of 70-80 nt, is a mixture of plant tRNA and CMV RNA fragments (Palukaitis et al., 1992). RNA 5 and RNA 6 each constitute 1-2% of total RNA in virus particles.
Protein: The capsid consists of 180 identical protein subunits of about Mr 24,500 (Van Regenmortel et al., 1972; Habili & Francki, 1974b). Coat protein can be isolated by disrupting virus particles and precipitating the RNA with 2M LiC1 (Francki et al., 1966). Coat protein sequences vary c. 2-3% within Subgroup II, 2-15% within Subgroup I, and c. 25% between subgroups I and II. Estimates of the coat protein Mr based on SDS-PAGE are often high, due to the arginine-rich, hydrophilic N-terminus (Hu & Ghabrial, 1995).
Other components: None reported.
Genome Properties
The CMV genome contains three messenger-sense RNAs designated RNA 1, RNA 2 and RNA 3 (Fig. 12). CMV-Fny RNA 1 (3,357 nt) encodes the c. 111 kDa 1a protein. CMV-Fny RNA 2 (3,050 nt) encodes the 97 kDa 2a protein, which is translated directly from RNA 2, and the 15 kDa 2b protein, which is translated from the 691 nt subgenomic RNA 4A. CMV-Fny RNA 3 encodes the 30 kDa movement protein, which is translated directly from RNA 3, and the 24.5 kDa coat protein, which is translated from the 1,034 nt subgenomic RNA 4 (Palukaitis et al., 1992; Ding et al., 1994). The 1a and 2a proteins plus host proteins comprise the CMV replicase (Hayes & Buck, 1990). The 2b protein is involved in inhibiting host responses restricting infection, and virus movement and is a virulence determinant (Ding et al., 1996; Li et al., 1999; Ji and Ding, 2001; Shi et al., 2002; Soards et al., 2002). The 3a protein is the viral movement protein (Kaplan et al., 1995). The 3b protein is the coat protein (Schwinghamer & Symons, 1975), and contains the determinants for aphid transmission (Mossop & Francki, 1977; Gera et al., 1979; Perry et al., 1994, 1998). Both the 3a protein and the coat protein are required for cell-to-cell movement as well as long-distance movement (Canto et al., 1997; Ryabov et al., 1999). The genetic mapping of these and several other phenotypic traits on the virus genome is given in Table 1.
Table 1. Genetic mapping of traits on the CMV genome
| RNA | Gene | Position | Phenotype | Reference |
| 1 | 1a? | NE a | Hypersensitive response in tobacco | Lakshman & Gonsalves, 1985 |
| 1 | 1a? | NE | Rapid local and systemic movement in squash | Gal-On et al., 1994 |
| 1 | 1a? | NE | Seed transmission in legumes | Hampton & Francki, 1992 |
| 1 | 1a | 173 | Replication (ts) b in melon | Bao et al., 1999 |
| 1 | 1a | 978 | Replication of satellite in squash | Roossinck et al., 1997 |
| 1 | 1a? | NE | Replication of satellite | McGarvey et al., 1995 |
| 2 | 2a | 631/641 | Hypersensitive response in cowpea | Kim & Palukaitis, 1997, Karasawa et al., 1999 |
| 2 | 2b | NE | Hypervirulence; systemic movement | Ding et al., 1995a; Brigneti et al., 1998; Shi et al., 2002; Soards et al., 2002 |
| 2 | 2b | NE | Suppression of gene silencing | Brigneti et al., 1998; Li et al., 1999 |
| 2 | NE | NE | Host range and/or pathology responses | Lakshman & Gonsalves, 1985; Palukaitis et al., 1992; Takeshita et al., 1998 |
| 3 | 3a | 20/21 | Limited movement between epidermal cells | Canto & Palukaitis, 1999a, 1999b |
| 3 | 3a | 20/21 | Tubule formation from protoplasts | Canto & Palukaitis, 1999b |
| 3 | 3a | 51 | Systemic movement in bottle gourd | Takeshita et al., 2001 |
| 3 | 3a | 51+240 | Systemic movement in cucurbits | Kaplan et al., 1997 |
| 3 | 3a | 60 | Systemic movement (ts) in tobacco | Li et al., 2001 |
| 3 | 3a | 97/98; 144/145; 156/158; 191/192c |
Cell-to-cell movement | Li et al., 2001 |
| 3 | 3a | 174-233 | RNA binding domain | Vaquero et al., 1997 |
| 3 | 3a | 247-279 | Capsid protein-dependent movementd | Nagano et al., 2001 |
| 3 | CP | 14-20 | Virion assembly | Schmitz & Rao, 1998 |
| 3 | CP | 26-40 | Virion assembly | Kaplan et al., 1998 |
| 3 | CP | 36 | Hypersensitive response on Nicotiana sp. | Takahashi et al., 2000 |
| 3 | CP | 81/166/173e | Virion assembly | Kaplan et al., 1998 |
| 3 | CP | 111/124 | Green mosaic on tobacco | Sugiyama et al., 2000 |
| 3 | CP | 129 | Chlorosis or veinal necrosis on tobacco | Shintaku et al., 1992; Suzuki et al., 1995 |
| 3 | CP | 129 | Virion assembly | Suzuki et al., 1995 |
| 3 | CP | 129 | Systemic movement in bottle gourd | Takeshita et al., 2001 |
| 3 | CP | 129 | Local symptom expression and systemic movement in Tetragonia expansa, Momordica charantia and Physalis floridana | Kobori et al., 2002 |
| 3 | CP | 129+162 | Local and systemic movement in maize | Ryu et al., 1998 |
| 3 | CP | 129+214 | Local and systemic movement in squash | Wong et al., 1999 |
| 3 | CP | 129+162+168 | Transmission by Aphis gossypii | Perry et al., 1994 |
| 3 | CP | 25+129+162+ 168+214 |
Transmission by Myzus persicae | Perry et al., 1994, 1998 |
| 3 | CP | 138 | Virion assembly | Suzuki et al., 1995 |
| 3 | CP | 144 | Virion assembly | Suzuki et al., 1995 |
| 3 | CP | 147 | Virion assembly | Suzuki et al., 1995 |
| 3 | CP | 162 | Virion stability | Ng et al., 2000 |
| 3 | CP | 193 | Stunting in Nicotiana glutinosa | Szilassy et al., 1999a |
| 3 | CP | NE | Systemic infection in Cucumis figarei | Saitoh et al., 1999 |
| 3 | RNA | 5' 618 nt | Necrotic etched ring in inoculated leaf | Zhang et al., 1994 |
| 3 | NE | NE | Host range and/or pathology responses | Palukaitis et al., 1992; Takeshita et al., 1998 |
a Not established.
b temperature sensitive.
c All four pairs of mutants individually affected virus movement.
d Deletion rendered cell-to-cell movement capsid-protein independent.
e Not delimited further.
The 3' termini of RNAs 1-4 are similar in sequence (Symons, 1979) and share a secondary structure that mimics that of tRNAs (Ahlquist et al., 1981). All four virus RNA species can be charged at the 3'-end with tyrosine by aminoacyl tRNA synthetases (Kohl & Hall, 1974). Their 5' ends are "capped" with 7-methyl guanosine (Symons, 1975). Ds RNA profiles from infected plants correspond to the three genomic RNA species and the two subgenomic RNA species, 4 and 4A.
The complete nucleotide sequences of at least 15 virus isolates and the coat protein gene sequences of over 50 isolates have been determined. The accession numbers and reference for the nucleotide sequences of the complete genome parts of various strains are:
Subgroup IA
| Strain Fny: | RNA-1, Accession D00356 (NC_002034) (Rizzo & Palukaitis, 1989); |
| RNA-2, Accession D00355 (NC_002035) (Rizzo & Palukaitis, 1988); | |
| RNA-3, Accession D10538 (NC_001440) (Owen et al., 1990). |
| Legume strain: | RNA-1, Accession D16403 (Karasawa et al., 1997); |
| RNA-2, Accession D16406 (Karasawa et al., 1997); | |
| RNA-3, Accession D16405 (Karasawa et al., 1997). |
| Strain Y: | RNA-1, Accession D12537 (Kataoka et al., 1990b); |
| RNA-2, Accession D12538 (Kataoka et al., 1990a); | |
| RNA-3, Accession D12499 (Nitta et al., 1988). |
| Ixora strain: | RNA-1, Accession U20220 (McGarvey et al., 1995); |
| RNA-2, Accession U20218 (McGarvey et al., 1995); | |
| RNA-3, Accession U20210 (McGarvey et al., 1995). |
| Strain NT9: | RNA-1, Accession D28778 (Hsu et al., 1995); |
| RNA-2, Accession D28779 (Hsu et al., 1995); | |
| RNA-3, Accession D28780 (Hsu et al., 1995). |
| Strain LS: | RNA-1, Accession AF416899 (Roossinck, 2002); |
| RNA-2, Accession AF416900 (Roossinck, 2002); | |
| RNA-3, Accession AF127976 (Roossinck et al., 1999). |
| Strain Q: | RNA-1, Accession X02733 (Rezaian et al., 1985); |
| RNA-2, Accession X00985 (Rezaian et al., 1984); | |
| RNA-3, Accession M21464 (Davies & Symons, 1988). |
| Strain Trk7: | RNA-1, Accession AJ007933 (Szilassy et al., 1999b); |
| RNA-2, Accession AJ007934 (Szilassy et al., 1999b); | |
| RNA-3, Accession L15336 (Salánki et al., 1994). |
Partial sequences from several other strains are also available in the database.
Satellite
Particles may contain small single stranded satellite RNA molecules (332-405 nt), which are not required for CMV replication, but that depend on CMV for replication, encapsidation and transmission (Kaper et al., 1976; Mossop & Francki, 1979).
When satellite RNA is present, up to 90% of the ds RNA present in infected plants may correspond to the satellite RNA, depending on the host, with a considerable reduction in the level of ds RNA 1 and RNA 2. The nucleotide sequences of over 40 satellite RNA isolates have been determined.
The secondary structure of some of satellite RNAs has been analysed in detail (Gordon & Symons, 1983, Bernal & García-Arenal, 1997; Rodríguez-Alvarado & Roossinck, 1997). The relationship between structure and function in CMV satellite RNAs has been investigated and models have been proposed (Palukaitis & García-Arenal, 1999).
Relations with Cells and Tissues
Virus particles can be difficult to detect in tissue sections by electron microscopy, because they are similar in size and appearance to ribosomes, although ribosomes can be digested without affecting the virus (Hatta & Francki, 1979). Particles, some of which appear hollow, are found scattered throughout the cytoplasm of infected cells. Particles can be identified in the cytoplasm, nuclei and vacuoles, but not in mitochondria, chloroplasts or plasmodesmata (Gerola et al., 1965; Lyons & Allen, 1969; Honda & Matsui, 1968, 1974; Ding et al., 1995b; Blackman et al., 1998). Aggregated particles bordered by a membrane structure are seen in phloem sieve elements (Blackman et al., 1998). Particles can also aggregate into crystalline arrays, usually in vacuoles (Martelli & Russo, 1985), where they can also be detected by light microscopy (Christie & Edwardson, 1977). In the cytoplasm, tubule-like structures composed of dictyosomal cisternae and containing virus-like particles can be seen late in infection (Martelli & Russo, 1985). Protoplasts expressing the jellyfish green fluorescent protein fused to the 3a protein, show fluorescent tubules extending into the suspension medium, in the presence or absence of coat protein (Canto & Palukaitis, 1999b); however, no such tubules are seen between any cell types in infected plants (Ding et al., 1995b; Blackman et al., 1998). The 3a protein is present in plasmodesmata between all leaf cell types (Blackman et al., 1998). The 2b protein is present in the nucleus (Mayers et al., 2000), and the 1a and 2a protein are membrane associated (Hayes & Buck, 1990), as is the replicase (Jaspars et al., 1985). Small membrane invaginations are sometimes seen associated with the tonoplast membrane (Hatta & Francki, 1981). Virus movement occurs through plasmodesmata, mediated by the 3a protein, with the coat protein being involved, but the formation of virions is not essential for cell-to-cell movement (Canto et al., 1997; Blackman et al., 1998; Kaplan et al., 1998). Coat protein is essential for long-distance movement (Ryabov et al., 1999).
Ecology and Control
In recent times CMV has caused severe epidemics in many crops, including necrosis of tomato in Italy (Gallitelli et al., 1988) (Fig. 13), Spain (Jordá et al., 1992) and Japan (Yoshida et al., 1984); mosaic and heart rot of banana world-wide (Bouhida & Lockhart, 1990; Singh et al., 1995); mosaic of melons in California (Grafton-Cardwell et al., 1996) and Spain (Luis-Arteaga et al., 1998); mosaic of borage in Spain (Luis-Arteaga et al., 1988); mosaic of pepper in Australia and California (Perry et al., 1993; Abdalla et al., 1991); and mosaic of lupins and other legumes in Australia and the USA (Jones & McLean, 1989; Jones & Coutts, 1997; Gillaspie et al., 1998). Economic losses, particularly in vegetable crops, can be very important (Tien & Wu, 1991, Jordá et al., 1992, Alonso-Prados et al., 1997). The effectiveness of control measures to eliminate reservoirs of virus inoculum depends largely on the ecological conditions, but is usually low because of the wide host range of CMV, its transmission by many aphid species, and its efficient transmission through seed in legumes and weeds. (Marchoux, 1990; Rist & Lorbeer, 1991). Insecticide treatments against aphid vectors is also of limited use (Marchoux, 1990; Bwye et al., 1997). Cross protection of plants with CMV strains supporting attenuating satellite RNAs has been an efficient method to control tomato necrosis in China and Italy (Tien & Wu, 1991; Crescenzi et al., 1993a). Genetic resistance to CMV is reported in many vegetable species and related wild relatives, but in most instances this resistance is strain-specific, and/or difficult to utilise in breeding programmes; resistance in commercial varieties exists only for cucumber (Havey, 1997). The Vat gene for resistance to CMV transmission by Aphis gossypii (Pitrat & Lecoq, 1980) is used in some commercial cultivars of melon. Transgenic resistance to CMV, associated with either coat protein transgenes, or truncated forms of the polymerase gene, is currently being evaluated in commercial varieties of cucurbits (Fuchs et al., 1998) and in advanced lines of tomato (Gal-On et al., 1998).
Notes
The physical, biological and antigenic properties of different cucumoviruses overlap making the use of these properties to distinguish them from particular isolates of CMV difficult. Combinations of serology and either nucleic acid hybridization (Wahyuni et al., 1992) or RT-PCR (Anonymous, 1998) have been used to distinguish isolates of CMV, and CMV from TAV and PSV. CMV isolates can also be distinguished by a ribonuclease protection assay (Fraile et al., 1997), which will also distinguish CMV from other viruses different in sequence from CMV. Alternatively, CMV can be distinguished from PSV and TAV by RT-PCR and restriction fragment length polymorphism, using a set of genus-specific primers and various restriction endonucleases (Choi et al., 1999). These various approaches should be more reliable than comparing differences in host range, and also can be used to distinguish CMV from other viruses with a broad host range and similar genome composition, such as Alfalfa mosaic virus.
Acknowledgements
We are grateful to Dr. M.Luis-Arteaga, SIA, Zaragoza, Spain for figures 1 to 10.
Figures

Large chlorotic local lesions observed in an inoculated leaf of sugarbeet (Beta vulgaris) caused by infected with strain Fny.

Pin-point necrotic local lesions in an inoculated leaf of Chenopodium amaranticolor.

Systemic chlorotic mottling in infected cucumber (Cucumis sativus cv. Marketer).

Systemic chlorotic mottling in infected squash (Cucurbita pepo cv, F1 Diamante).

Reduction in leaf size and shape in tomato (Lycopersicon esculentum cv. Rutgers) (right) systemically infected with strain WL (without satellite RNA). Uninfected plant on the left.

Systemic chlorotic mosaic with blisters in tobacco (Nicotiana tabacum cv. Paraguay) induced by infection with a Subgroup IA isolate.

Systemic yellow chlorosis in tobacco (Nicotiana tabacum cv. Xanthi nc) induced by infection with strain M.

Local necrotic lesions and etched rings in a leaf of tobacco (Nicotiana tabacum cv. Xanthi nc) induced by inoculation with a Subgroup II isolate.
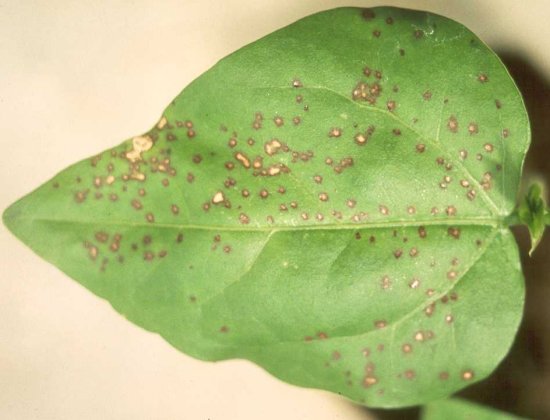
Large necrotic lesions in a leaf of cowpea (Vigna unguiculata cv. Blackeye 5) induced by inoculation with a Subgroup IA isolate.

Pin-point necrotic local lesions in a leaf of cowpea (Vigna unguiculata cv. Blackeye 5) induced by inoculation with a Subgroup II isolate.
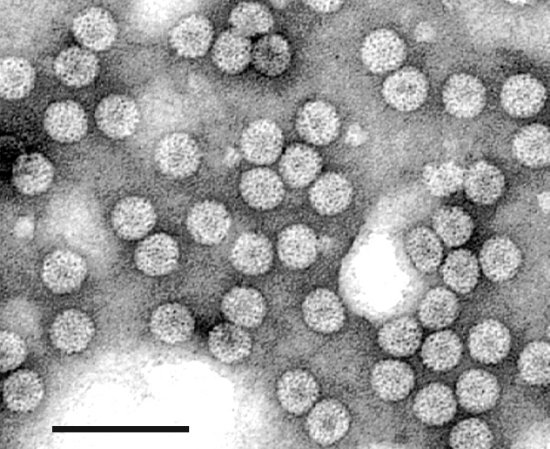
Electron micrograph of a purified preparation of virus particles stained with uranyl acetate. Bar represents 100 nm.
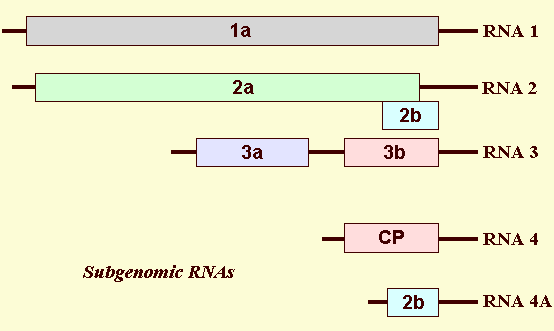
The genome organisation of Cucumber mosaic virus. The three genomic RNAs (1, 2 and 3), as well as the two subgenomic RNAs (RNAs 4 and 4A), are shown. RNA 1 encodes the 1a protein. RNA 2 encodes the 2a protein and the 2b protein, but the 2b protein is only expressed from the subgenomic RNA 4A. RNA 3 encodes the 3a and capsid protein (CP), but the CP is only expressed from the subgenomic RNA 4. Subgenomic RNA 4 is encapsidated in virions. Subgenomic RNA 4A is only encapsidated into virions of subgroup II CMV strains.

A field of tomato (Lycopersicon esculentum) plants in the Puglia district of southwest Italy infected with CMV containing a necrosis-inducing satellite RNA. Unlike plants infected within a few weeks after germination, plants infected later show necrosis on stems and leaves, but not always lethal necrosis.