Details of DPV and References
DPV NO: 407 December 2004
Family: Secoviridae
Genus: Waikavirus
Species: Rice tungro spherical virus | Acronym: RTSV
This is a revised version of DPV 67
Rice tungro spherical virus
Roger Hull Department of Disease and Stress Biology, John Innes Centre, Norwich Research Park, Colney, Norwich NR4 7UH, UK
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Rice tungro disease was first described by Anon. (1964) and Rivera & Ou (1965). The first description of rice tungro spherical virus (RTSV) was by Galvez (1968) and the role of RTSV as a member of the virus complex that causes tungro disease was recognized by Hibino et al. (1978). The cause of waika disease of rice, first described by Saito (1977), was reported by Anjaneyulu et al. (1995).
- Synonym
- Rice waika virus (Saito, 1977).
A virus with isometric particles about 30 nm in diameter, which contain a single molecule of positive-sense ssRNA of over 12 kb, and three proteins, CP1, CP2 and CP3, of Mr 22,900, 22,300 and 33,000, respectively. The virus is transmitted in a semi-persistent manner by leafhoppers; it is not mechanically transmissible. It occurs in Oryza species in South and South-East Asia. It acts as a helper for leafhopper transmission of Rice tungro bacilliform virus (RTBV). In most cultivars of rice, RTSV causes no symptoms but in some cultivars it causes a mild stunting disease (waika); the RTSV/RTBV complex causes a severe disease (tungro).
Main Diseases
In most rice cultivars, infection with RTSV alone results in almost no symptoms and the plants appear green with mild or no stunting (Hibino et al., 1978). In some rice cultivars, e.g. Reiho (Norin 100), the virus causes noticeable stunting (waika) (Furuta, 1977). RTSV in complex with Rice tungro bacilliform virus (RTBV) induces the economically important rice tungro disease (Hibino et al., 1978). Annual losses due to tungro disease in South-East Asia have been estimated to exceed $1.5 billion (Herdt, 1991).
Geographical Distribution
The virus is found in most, if not all, rice-growing countries of South and South-East Asia. Because it usually causes symptomless infections, it is probably more widespread than is currently recognized. Its distribution is likely to be related to that of its major leafhopper vectors.
Host Range and Symptomatology
Early host range studies on RTSV are unreliable as its association with RTBV was not then recognized. Host range studies are also constrained by the ability of the vector species to transmit the virus to the plant species tested. RTSV appears to have a restricted host range limited to members of the Poaceae and Cyperaceae (Echinochloa crus-galli, E. glabescens, E. colona, Leptochloa chinensis, Leersia hexandra, Oryza sativa, Panicum repens, Cyperus rotundus) (Khan et al., 1991). RTSV causes few, if any, symptoms in these hosts.
Because of lack of symptoms, the most reliable diagnosis is by serological or nucleic acid-based techniques. The virus can be propagated in rice cultivar TN1.
Strains
The type strain of RTSV can be considered to be the Philippines isolate that was the first sequenced (Shen et al., 1993). RTSV strain Vt6 overcomes the resistance to the type strain in rice cultivar TKM6 (Cabauatan et al., 1995). Several serological variants of RTSV have been reported (Druka et al., 1996; Druka & Hull, 1998). Molecular techniques reveal much microvariation in the sequences encoding CP1 and CP2 (Azzam et al., 2000).
Transmission by Vectors
RTSV is transmitted in the semi-persistent manner by several species of leafhopper. The major vector over much of South-East Asia is the rice green leafhopper, Nephotettix virescens (synonym N. impicticeps) (Fig. 1); the virus is also transmitted by N. cincticeps, N. nigropictus, N. malayanus and Recilia dorsalis. These other vectors may be of relative importance in certain situations. N. virescens can acquire the virus in 30 min and there is no latent period; it can retain infectivity for up to 4 days (Anjaneyulu et al., 1995). All five larval stages and adults transmit the virus (Ling, 1966) but nymphs transmit more efficiently than adults and females more efficiently than males. Transmission efficiency is also affected by temperature, increasing as the temperature rises from 10 to 34 °C.
Transmission through Seed
No evidence for seed transmission.
Serology
RTSV is moderately immunogenic and the antisera have been used for ELISA, western blots and immunogold electron microscopy (Bajet et al., 1985; Cabauatan & Hibino, 1988; Druka et al., 1996). Antisera have also been raised to the three individual coat protein species expressed in E. coli and it was shown that CP3 is the major antigenic determinant on the surface of the virus particle (Druka et al., 1996).
Relationships
The physical and chemical properties of RTSV are similar to those of other members of the Sequiviridae but no relationships have been shown between RTSV and other viruses. Strains of RTSV can be differentiated by ability to infect rice cultivar TKM6, and isolates can be differentiated by the serological behaviour of CP3 (Druka et al., 1996).
Stability in Sap
As the virus is not sap-transmissible, there have been no studies on its stability in sap.
Purification
1. Method of Cabauatan & Hibino (1988):
Harvest rice plants 50 days after infection and homogenize each 500 g material in 1 L 0.01 M sodium
ethylene-diamine-tetra-acetate (EDTA), pH 8.0. Incubate the extract with 3 g Driselase
(Kyowa Hakko Kogyo Co. Ltd., Tokyo) for 1 h at room temperature,1 h at 40 °C, and then centrifuge for 10 min
at 15,000g. Add polyethylene glycol (mol. wt 8000), NaCl and Triton X-100 to 7%, 0.2 M
and 1% respectively and stir the mixture for 1 h at room temperature, centrifuge at 30,000g
for 30 min and resuspend the pellets in 20 ml cold 0.01 M EDTA, pH 8.0. Centrifuge at 11,000g
for 10 min, retain the supernatant fluid and centrifuge it at 100,000g for 60 min.
Resuspend the pellet in 2 ml cold 0.01 M phosphate buffer (PB), pH 7.4, layer on 10-50% sucrose density gradients
(in PB) and centrifuge in a Beckman SW27 rotor at 25,000 rev./min for 2.5 h at 4 °C. Recover the
virus-containing band, dilute in 0.01 M PB and centrifuge at 130,000g for 1 h. Resuspend the
resulting pellet in 1 ml PB and centrifuge at 11,000g for 10 min. The virus is found in the
supernatant fluid.
2. Method of Jones et al. (1991):
Freeze infected rice leaves in liquid N2, grind and thaw each 100 g tissue into 400 ml 0.1 M
sodium citrate, pH 5.9. Add Celluclast (Novo Enzymes) to 5% (v/v), incubate the mixture at 30 °C for 2 h
and centrifuge at 10,000 rev./min for 10 min. Resuspend the pellet in 200 ml citrate buffer
and 5% Celluclast, incubate for 1 h at 30 °C and centrifuge as above. Pool the supernatant fractions from
both centrifugations and add polyethylene glycol (mol. wt 6000), NaCl and Triton X-100 to give 7%, 0.2 M
and 1% respectively. Incubate the mixture for 2 h at room temperature, then centrifuge at 12,000 rev./min
for 10 min. Resuspend the pellet in 20 ml citrate buffer and centrifuge through a 5% sucrose cushion
at 36,000 rev./min for 2.5 h in a Beckman Ti40 rotor at 4 °C. Resuspend the pellet in 4 ml citrate buffer.
Further purification and, if necessary, separation from RTBV can be effected by rate zonal or isopycnic gradient
centrifugation.
Properties of Particles
The particles sediment with an so20,w in the range of 150-190 S. Other physical properties have not been determined.
Particle Structure
The virus particles are isometric, about 30 nm in diameter (Fig. 2). The three coat proteins are in apparently equimolar amounts (Hull, 1996) and, by analogy with comoviruses and picornaviruses, the structure is considered to be a T = 1 icosahedron.
Particle Composition
Nucleic acid:
Virus particles contain one species of positive-sense ssRNA. The complete nucleotide sequence has been
determined for the type isolate (12,226 nucleotides;
accession number NC001632)
(Shen et al., 1993;
Thole & Hull, 1996)
and for the resistance-breaking strain, Vt6 (12,171 nucleotides;
accession number AB064963)
(Isogal et al., 2000).
Protein:
Polyacrylamide gel electrophoresis of viral coat protein reveals three species, CP1, CP2 and CP3, of 21.5, 24
and 31 kDa
(Hibino et al., 1991)
or 25, 26 and 35 kDa
(Jones et al., 1991),
respectively. Identification of cleavage sites by N-terminal sequencing shows that CP1 is 22.9 kDa and
CP2 is 22.3 kDa
(Shen et al., 1993;
Zhang et al., 1993);
the C-terminus of CP3 has not been identified. In western blots of crude extracts from infected plants,
an antiserum to CP3 identifies a band at 33 kDa and several in the range 40-42 kDa; the nature of these
larger bands is unknown
(Druka et al., 1996).
Genome Properties
The genome encodes a polyprotein of 393 kDa (Fig. 3) and has two possible small open reading frames (ORFs) at the 3' end (Shen et al., 1993); however, the nature of these 3' ORFs has been questioned (Thole & Hull, 1996).
The cleavage sites of two of the three coat protein species (CP1 and CP2) and of the C-terminal cysteine protease and polymerase products have been determined (Shen et al., 1993; Zhang et al., 1993; Thole & Hull, 1998). Little is known of the products N-terminal to the coat proteins or in the region downstream of CP3. An antiserum to E. coli-expressed CP1 detected a product of 32 kDa, much smaller than the expected 70 kDa (Hull, 1996); the region downstream of CP3 contains the sequence motif for a nucleotide triphosphate binding protein (Shen et al., 1993).
Relations with Cells and Tissues
RTSV is limited to vascular tissues and is restricted to the phloem cells (Sta. Cruz et al., 1993). Virus particles are scattered in the cytoplasm (Fig. 4) or are embedded in non-enveloped electron-dense granular inclusions (Fig. 5). Particles also occurred as crystalline aggregates, often in vacuoles (Fig. 6). Small vesicles containing fibres are found in the cytoplasm, usually along the wall of infected cells (Fig. 7).
Ecology and Control
RTSV on its own causes no significant damage in most rice cultivars. However, it provides the transmission capability to RTBV in the economically important tungro complex and thus has considerable importance in relation to that disease. It spreads from old to seedling plants in overlapping cropping systems and is likely to spread from latent infections in weeds harbouring the leafhopper, Nephotettix virescens, surrounding rice paddies. Tungro infection occurs in periodic epidemics (Savary et al., 1993; Anjaneyulu et al., 1995).
Control of RTSV is either by use of insecticides (see Anjaneyulu et al., 1995) or by resistance breeding. Rice breeding programmes have attempted to introduce resistance to RTSV into elite cultivars. Resistance to the vector is not very durable (Dahal et al., 1990). Resistance to the virus occurs in varieties TKM6 (Hibino et al., 1988) and TW5 (Azzam et al., 2001), that of TKM6 co-segregating with leafhopper resistance (Sebastian et al., 1996); however, RTSV strain Vt6 breaks this resistance (Cabauatan et al., 1995). A single recessive gene controls the TKM6 resistance, whereas in TW5 two recessive genes are involved (Azzam et al., 2001). Transformation of rice with coat protein or replicase genes affords protection against the virus (Sivamani et al., 1999; Huet et al., 1999).
Figures

The rice green leafhopper, Nephotettix virescens.
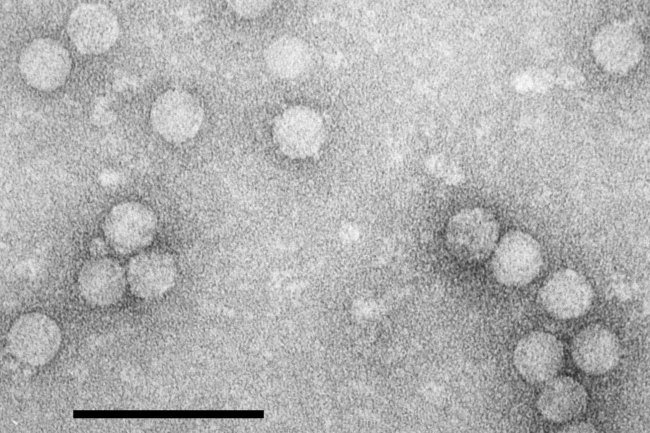
Particles of RTSV negatively stained in uranyl acetate. Bar represents 100 nm.
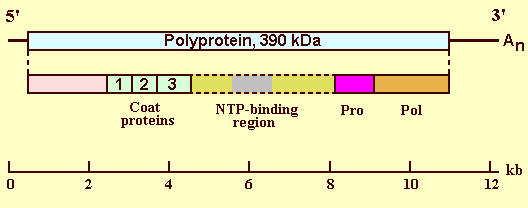
Genome organization of RTSV. The horizontal line represents the genomic positive-sense ssRNA with the 3' polyadenylation (An) indicated. The box shows the polyprotein with below the positions of the processed coat proteins (CP1, CP2 and CP3), protease (Pro) and polymerase (Pol) indicated together with the motif for the NTP binding site.

Thin sections of rice plant, cultivar TN1, jointly infected with RTSV and RTBV showing both viruses in the cytoplasm. Bar represents 400 nm. (From Sta. Cruz et al., 1993).

Thin sections of rice plant, cultivar TN1, jointly infected with RTSV and RTBV showing RTSV embedded in an inclusion body in the cytoplasm. Bar represents 400 nm. (From Sta. Cruz et al., 1993).

Thin sections of rice plant, cultivar TN1, jointly infected with RTSV and RTBV showing RTSV as crystalline aggregates. Bar represents 400 nm. (From Sta. Cruz et al., 1993).

Thin sections of rice plant, cultivar TN1, jointly infected with RTSV and RTBV showing vesicles (VE) containing fibres (F) and RTSV particles. Bar represents 400 nm. (From Sta. Cruz et al., 1993).
References list for DPV: Rice tungro spherical virus (407)
- Anjaneyulu, Satapathy & Shukla, Rice tungro. Science Publishers, Inc., Lebanon, New Hampshire, 228 pp., 1995.
- Anon., Report of the International Rice Research Institute, 1963, 1964.
- Azzam, Yambao, Muhsin, McNally & Umadhay, Archives of Virology 145: 1183, 2000.
- Azzam, Imbe, Ikeda, Nath & Coloquio, Euphytica 122: 91, 2001.
- Bajet, Daquioag & Hibino, Journal of Plant Protection in the Tropics 2: 125, 1985.
- Cabauatan & Hibino, Plant Disease 72: 526, 1988.
- Cabauatan, Cabunagan & Koganezawa, Phytopathology 85: 77, 1995.
- Dahal, Hibino & Saxena, Phytopathology 80: 371, 1990.
- Druka & Hull, Journal of Phytopathology 146: 175, 1998.
- Druka, Burns, Zhang & Hull, Journal of General Virology 77: 1975, 1996.
- Furuta, Review of Plant Protection Research 10: 70, 1977.
- Galvez, Virology 35: 418, 1968.
- Herdt, Rice Biotechnology, Eds Khush & Toenniessen, pp. 19-54, CABI, Wallingford, UK, 1991.
- Hibino, Roechan & Sudarisman, Phytopathology 68: 1412, 1978.
- Hibino, Daquioag, Cabauatan & Dahal, Plant Disease 72: 843, 1988.
- Hibino, Ishikawa, Omura, Cabauatan & Koganezawa, Phytopathology 81: 1130, 1991.
- Huet, Mahendra, Wang, Sivamani, Ong, Chen, de Kochko, Beachy & Fauquet, Phytopathology 89: 1022, 1999.
- Hull, Annual Review of Phytopathology 34: 275, 1996.
- Isogal, Cabauatan, Masuta, Uyeda & Azzam, Virus Genes 20: 79, 2000.
- Jones, Gough, Dasgupta, Subba Rao, Cliffe, Qu, Shen, Kaniewska, Blakebrough, Davies, Beachy & Hull, Journal of General Virology 72: 757, 1991.
- Khan, Hibino, Aguiero & Daquioag, Plant Disease 75: 926, 1991.
- Ling, Phytopathology 56: 1252, 1966.
- Rivera & Ou, Plant Disease Reporter 49: 127, 1965.
- Saito, Review of Plant Protection Research 10: 83, 1977.
- Savary, Fabellar, Tiongco & Teng, Plant Disease 77: 376, 1993.
- Sebastian, Ikeda, Huang, Imbe, Coffman & McCouch, Phytopathology 86: 25, 1996.
- Shen, Kaniewska, Smith & Beachy, Virology 193: 621, 1993.
- Sivamani, Huet, Shen, Ong, de Kochko, Fauquet & Beachy, Molecular Breeding 5: 177, 1999.
- Sta. Cruz, Koganezawa & Hibino, Journal of Phytopathology 138: 274, 1993.
- Thole & Hull, Virus Genes 13: 239, 1996.
- Thole & Hull, Virology 247: 106, 1998.
- Zhang, Jones, Barker, Davies & Hull, Virus Genes 7: 121, 1993.