Details of DPV and References
DPV NO: 409 June 2005
Family: Partitiviridae
Genus: Alphacryptovirus
Species: White clover cryptic virus 1 | Acronym: WCCV-1
White clover cryptic virus 1
Robert G. Milne Istituto di Virologia vegetale, CNR, 10135 Torino, Italy
Thierry Candresse Equipe de Virologie, UMR GD2P, INRA et Université Bordeaux 2, IBVM, Campus INRA, BP81, 33883 Villenave d'Ornon Cedex, France
Guido Boccardo Istituto di Virologia Vegetale, CNR, 10135 Torino, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Boccardo et al. (1985).
A virus with isometric particles about 30 nm in diameter containing two dsRNA species of estimated sizes 2.0 and 1.7 kbp and a major coat protein of Mr 55,000. Occurs in white clover (Trifolium repens) probably worldwide, but causes no symptoms. Not transmissible by sap inoculation or by grafting but transmitted to a high degree through the seed. No vector is known.
Main Diseases
Causes no symptoms in white clover, the only host known (Boccardo et al., 1985, 1987).
Geographical Distribution
Found in white clover grown from seed of all cultivars from Europe, New Zealand and the USA that were tested but not in any cultivars from South Australia (Boccardo et al., 1985); found in cv 'California' grown in Japan (Natsuaki et al., 1986).
Host Range and Symptomatology
Probably confined to Trifolium repens (white clover). The virus was found in seed samples from 8 of 13 cultivars (Boccardo et al., 1985; Natsuaki et al., 1986). The experimental host range has not been determined because of the difficulty of inter-species transmission.
- Diagnostic species
- No diagnostic species known.
- Propagation species
- Any white clover plant carrying the virus, vegetatively propagated, and screened for the absence of other viruses, in particular other cryptic viruses. Presence of the virus in vegetatively propagated carrier clones must be confirmed periodically because virus-free stolons sometimes arise (see Relations with Cells and Tissues).
- Assay species
- No assay species known.
Strains
No differences detected, using DAS ELISA with antiserum to one isolate.
Transmission by Vectors
Not tested; probably not vector-transmissible.
Transmission through Seed
The percentage of white clover seedlings carrying WCCV-1 ranged from 35 to 74% in seven cultivars (Boccardo et al., 1985), but these tests were made on commercial seed, and the status of the parent plants was unknown. Of several other cryptic viruses tested, all were transmitted to the seed via pollen as well as via the female parent (Boccardo et al., 1987). Thus with Beet cryptic virus (BCV; Kassanis et al., 1978) the rate of transmission via the ovum was 82%, and that via the pollen was 43%; if both parents were carriers, 100% of the seed gave rise to carrier seedlings. Similar results were obtained with Vicia cryptic virus (Kenten et al., 1980). Antoniw et al., (1990) used transmission rates for BCV to model the spread of cryptic viruses through a population of 100,000 plants starting with a single carrier plant. The model predicted that almost the entire population would become carriers after about 80 generations. A similar outcome seems likely for WCCV-1.
Transmission by Grafting
No transmission (Boccardo et al., 1985, 1987).
Transmission by Dodder
Not transmitted (V. Lisa & E. Luisoni, unpublished).
Serology
The virus is a good immunogen and antigen. Antisera were obtained with homologous gel-diffusion titres of 1/2048 and EM decoration titres of 1/4096, with titres of 1/4 against preparations of dsRNA. The most effective serological tests have been gel double diffusion (Fig. 1), DAS ELISA, ISEM and EM decoration (Fig. 2) (Boccardo et al., 1985; Natsuaki et al., 1986).
Relationships
Antisera to WCCV-1 did not react with any other cryptic virus or mycovirus tested (Boccardo et al., 1985; Luisoni et al., 1987).
Stability in Sap
No information.
Purification
Use T. repens plants known to be carrying WCCV-1 only. Operate at 4 °C. Homogenise 1 kg of leaves and stems with 2.5 litres of 50 mM potassium phosphate buffer, pH7, containing 20 mM Na2SO3, 10 mM DIECA and 5 mM EDTA. Emulsify with an equal volume of chloroform and after separation of the emulsion dissolve 10% (w/v) PEG (MW 6000) and 1% (w/v) NaCl in the aqueous phase. After stirring for 2.5 h, centrifuge at 13,700 g for 10 min and resuspend the pellets in 450 ml of 500 mM phosphate buffer, pH 7 containing 20 mM Na2SO3. Treat the suspension with chloroform as above, then ultracentrifuge the aqueous phase at 300,000 g for 60 min. Resuspend the pellets in 15 ml of 50 mM phosphate buffer, pH 7, and clarify by low-speed centrifugation. Layer the supernatant on preformed 25-45 % gradients of Cs2SO4 prepared in the same buffer and centrifuge for 60 min at 36,000 rpm in a Beckman SW 41Ti rotor or equivalent. Extract the virus-rich band, seen by light-scattering (Fig. 3) and identified by electron microscopy (Fig. 4), dilute in buffer, and resediment the virus particles by ultracentrifugation at 300,000 g for 60 min. The yield is about 150 µg of virus/kg of tissue. For alternatives, see Lisa et al. (1981), Boccardo et al. (1985) and Boccardo et al. (1987).
Properties of Particles
After isopycnic centrifugation in CsCl, the virus forms one band with a buoyant density of 1.392 g/ml. The A260/A280 ratio is 1.37, and Amax/Amin is 1.19 (values uncorrected for light-scattering); the particles are stable in neutral 10 mM phosphate buffer for many weeks at 4 °C (Boccardo et al., 1985).
Particle Structure
The particles (Fig. 4) when negatively stained in 2% (w/v) uranyl acetate, appear about 30 nm in diameter, rounded in profile and without visible substructure. Boccardo et al. (1985) estimated the particle diameter as 34 nm but this was later corrected to 29-32 nm (Boccardo et al., 1987) and 30 nm (Ghabrial et al., 2000).
Particle Composition
The particles contain 2 dsRNAs estimated by gel electrophoresis to be 2.0 and 1.7 kbp in size (Fig. 5) (Boccardo et al., 1985; Luisoni et al., 1987). There is one major capsid protein estimated to be 55 kDa in size, and RNA polymerase activity is associated with the particles (Boccardo & Accotto, 1988).
Genome Properties
The two genomic dsRNAs have been sequenced (accession numbers AY705784and AY705785; Boccardo et al., 1990, 1992; Boccardo & Candresse, 2005a, 2005b). Each RNA contains a single open reading frame, and each coding strand is polyadenylated at the 3' end (Fig 6). RNA1 has a size of 1955 nt excluding the poly-A tail, and a G+C content of 44%. RNA2 is 1708 nt in size, excluding the poly-A tail, with a G+C content of 49.6%. The 5' non-coding regions are respectively 74 (RNA1) and 104 (RNA2) nt long and are about 76% identical. By contrast the two 3' NCRs are very different in size (30 nt, 140 nt) and show no sequence homology.
RNA1 encodes a protein of 616 amino acids with a calculated Mr of 72.9 kDa; the protein contains the GDD motif typical of RNA-dependent RNA polymerases (RdRPs) and has homologies with the RdRPs of other members of the Partitiviridae. RNA2 encodes a protein of 487 amino acids with a calculated Mr of 54.2 kDa, confirmed as the coat protein by in vitro translation and immunoprecipitation experiments (Boccardo & Candresse, 2005b). Apparently the genome encodes only these two proteins.
Relations with Cells and Tissues
The virus occurs in all tissues examined: ovaries, stamens, petals, leaves, roots, epidermal strips from stolons, and stolon segments with the epidermis removed (Boccardo et al., 1985).
A carrier seedling of T. repens initially has the virus in all tissues but when creeping stolons develop, some of these become virus-free (Boccardo et al., 1987).
Ecology and Control
The virus is probably widespread in T. repens, being transmitted through pollen and ovules to a high percentage of seed, but there appear to be no vectors. Since no damage is caused by the virus, control has not been attempted.
Notes
WCCV-1 has been designated as the type species of the genus Alphacryptovirus, differentiated from a second genus of plant cryptic viruses, Betacryptovirus (type species White clover cryptic virus 2, WCCV-2) (Luisoni & Milne, 1988). These two genera fall within the family Partitiviridae, otherwise made up of viruses symptomlessly infecting fungi. The distinction between alpha and beta cryptoviruses so far rests only upon the distinct virus particle morphologies (betacryptovirus particles are larger, with distinct subunits), since there are no data available on sequences or coat protein sizes for any betacryptovirus.
Some sequence data exist for a number of alphacryptoviruses: Alfalfa cryptic virus (Accotto et al., 1990), Beet cryptic virus 1 (Accotto et al., 1987), Beet cryptic virus 3 (S63913; Xie et al., 1993), Vicia cryptic virus (Y09237) and a cryptic virus from Pyrus pyrifolia (AB012616; Osaki et al., 1998). Homology comparisons of the available polymerase gene sequences (Boccardo & Candresse, 2005a) indicate that WCCV-1 is related to the alphacryptovirus Vicia cryptic virus (79% identity with the partial VCV sequence available) and to a member of the Partitivirus genus of mycoviruses (close to 60% identity with the polymerase gene of Heterobasidion annosum virus, AF348136). However, WCCV-1 is only distantly related to a second alphacryptovirus, Beet cryptic virus 3 (about 20% identity between the polymerase amino acid sequences) whereas the BCV-3 polymerase shows about 53% amino acid sequence identity with that of the cryptic virus in Pyrus pyrifolia. This suggests that the polymerases of alphacryptoviruses may be polyphyletic in origin, and that the taxonomy of plant cryptic viruses will need re-examination.
Figures
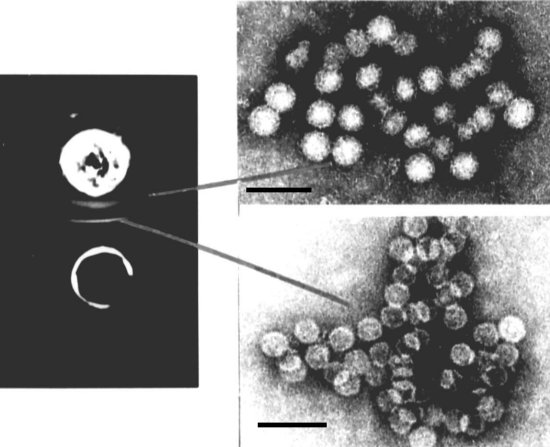
Agar gel double-immunodiffusion test using a natural mixture of white clover cryptic viruses 1 and 2 (upper well), and antiserum to the mixed viruses (lower well). Two bands were formed, identified by electron microscopy as composed of aggregated particles of WCCV-2 (upper right) and WCCV-1 (lower right). Bars = 100 nm. (courtesy of E. Luisoni and R.G. Milne).
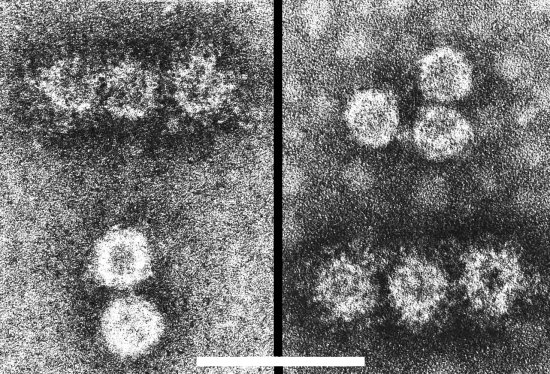
Particles of WCCC-1 and WCCV-2 from crude sap of a white clover plant were trapped on an EM grid coated with an antiserum reacting to both particles. The grid was then further decorated with antiserum to WCCV-1 (left panel) or WCCV-2 (right panel). Bar = 100 nm. (Natsuaki et al., 1986).
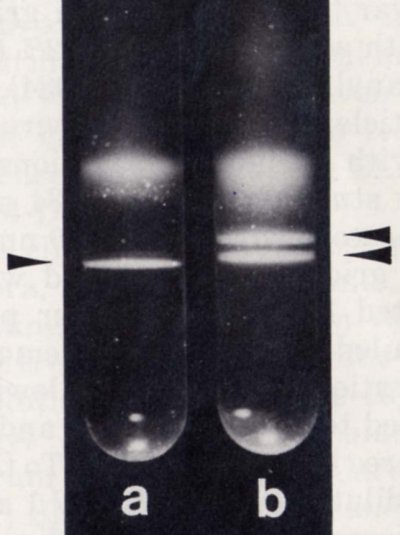
Light-scattering bands formed after purified virus preparations were centrifuged into preformed caesium sulphate density gradients. a) WCCV-1; b) mixture of WCCV-1 and WCCV-2 (Boccardo et al, 1985).

Purified preparation of WCCV-1 particles negatively stained in uranyl acetate. Bar = 100 nm.

Electrophoretic separation in a 5% polyacrylamide slab gel of dsRNA from WCCV-1 alone (A), WCCV-1 + WCCV-2 (B), WCCV-2 alone (C), Maize rough dwarf virus (D) and a cypovirus (E; gift of C. C. Payne). Stained with silver.
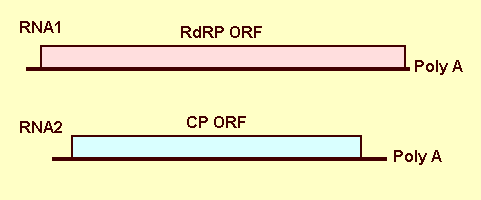
Genomic organization of WCCV-1, showing the two RNAs, RNA1 encoding the polymerase, RNA 2 encoding the coat protein.
References list for DPV: White clover cryptic virus 1 (409)
- Accotto, Brisco & Hull, Journal of General Virology 68: 1417, 1987.
- Accotto, Marzachí, Luisoni & Milne, Journal of General Virology 71: 433, 1990.
- Antoniw, White & Xie, in Recognition and Response in Plant-Virus Interactions, p. 273, ed. R. S. S. Fraser, Berlin: Springer-Verlag, 1990.
- Boccardo & Accotto, Virology 163: 413, 1988.
- Boccardo & Candresse, Archives of Virology 150: 399, 2005a.
- Boccardo & Candresse, Archives of Virology 150: 403, 2005b.
- Boccardo, Milne, Luisoni, Lisa & Accotto, Virology 147: 29, 1985.
- Boccardo, Lisa, Luisoni & Milne, Advances in Virus Research 32: 171, 1987.
- Boccardo, Candresse, d'Aquilio & Dunez, Abstracts, VIIIth International Congress of Virology, W88-005, p. 133, 1990.
- Boccardo, Candresse, d'Aquilio & Dunez, Abstracts, 4th International Symposium on Double-Stranded RNA Viruses, S7-8, p. 35, 1992.
- Ghabrial, Buck, Bozarth, Martelli & Milne, in Virus Taxonomy, Seventh Report of the International Committee on Taxonomy of Viruses, p. 508, ed. M. V. H. Van Regenmortel et al., New York, San Diego: Academic Press, 2000.
- Kassanis, Russell & White, Phytopathologische Zeitschrift 91: 76, 1978.
- Kenten, Cockbain & Woods, Rothamsted Experimental Station Report for 1979, Part 1, p. 176, 1980.
- Lisa, Luisoni & Milne, Annals of Applied Biology 98: 431, 1981.
- Luisoni & Milne, AAB Descriptions of Plant Viruses 332, 1988.
- Luisoni, Milne, Accotto & Boccardo, Intervirology 28: 144, 1987.
- Natsuaki, Natsuaki, Okuda, Teranaka, Milne, Boccardo & Luisoni, Intervirology 25: 69, 1986.
- Osaki, Kudo & Ohtsu, Bioscience, Biotechnology and Biochemistry 62: 2101, 1998.
- Xie, Antoniw & White, Journal of General Virology 74: 1467, 1993.