Details of DPV and References
DPV NO: 417 July 2007
Family: Caulimoviridae
Genus: Petuvirus
Species: Petunia vein clearing virus | Acronym: PVCV
Petunia vein clearing virus
Katja R. Richert-Pöggeler Federal Biological Research Centre, Dept. of Plant Virology, Microbiology and Biosafety, Messeweg 11-12, D-38104 Braunschweig, Germany
Dietrich-E. Lesemann Federal Biological Research Centre, Dept. of Plant Virology, Microbiology and Biosafety, Messeweg 11-12, D-38104 Braunschweig, Germany
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Lesemann & Casper (1973).
Petunia vein clearing virus has a double-stranded (ds) DNA genome encapsidated by isometric virions (Fig. 1), slightly smaller than those of Cauliflower mosaic virus (CaMV). Inclusion bodies similar to those described for CaMV can be seen in the cytoplasm of infected cells (Fig. 2). It is the type member of the genus Petuvirus. Virus infection can arise from PVCV sequences integrated into the host genome. Endogenous full-length copies of PVCV can be induced by wounding and have been shown to be infectious (Richert-Pöggeler et al., 2003). Horizontal transmission of the virus is possible via grafting. The virus only infects solanaceous hosts and occurs worldwide (Harper et al., 2003; Lockhart & Lesemann, 1997; Zeidan et al., 2001).
Main Diseases
The virus was first reported in Petunia × hybrida hort. cv. Himmelsroeschen in Germany (Lesemann & Casper, 1973). The only known naturally-occurring disease caused by PVCV is in members of the genus Petunia, including commercially-available hybrid petunia (Harper et al., 2002; Staginnus & Richert-Pöggeler, 2006). Symptoms consist of vein clearing of the major leaf veins (Fig. 3). Symptoms are rarely seen under standard growing conditions but abiotic stresses, like water deficiency, can trigger viral infection (Lockhart & Lesemann, 1997). In addition, wounding and elevated temperatures can also induce symptom expression (Richert-Pöggeler et al., 2003; Staginnus & Richert-Pöggeler, 2006; Noreen et al., 2007). Mixed infections with Potato virus Y, Tobacco mosaic virus and Cucumber mosaic virus have been observed (Richert, 1992).
Geographical Distribution
Using immunosorbent electron microscopy, PVCV-infected petunia plants have been identified worldwide, including the USA, Israel, the United Kingdom, the Netherlands, Germany and Japan (Lesemann, unpublished).
Host Range and Symptomatology
Characteristic vein-clearing symptoms (Fig. 4) occur 6-8 weeks after grafting of infected petunia scions or after biolistic inoculation using recombinant PVCV constructs (Richert-Pöggeler et al., 2003). The virus infects the plant systemically. Symptoms appear in newly developed leaves and young (side) shoots. The chlorosis along the veins can be of different shapes: i) restricted to the leaf vein, ii) starting from the midrib and spreading in a more diffuse pattern to adjacent leaf parts and iii) occurring as spots along the major veins (Fig. 3). All three types of leaf symptom can occur on the same plant. In the original host, cv. Himmelsroeschen, chlorosis has also been observed on stem parts (Lesemann & Casper, 1973). Additionally, heavily infected plants can display a disturbed colour pattern in their flowers (Richert, 1992) and leaf malformation (Lesemann, 2006). Epinasty sometimes occurs on symptomatic leaves (Fig. 4).
The virus has a limited host range, infecting only a small number of species from the family Solanaceae. Besides various petunia species (P. axillaris, P. integrifolia and P. hybrida), the virus could be transmitted by grafting to Nicotiana glutinosa and N. benthamiana; plants that do not carry any endogenous PVCV sequences in their genomes. Symptom expression in Nicotiana spp. consists of vein-clearing, but the chlorosis is less extensive than in Petunia spp., and covers only segments within the main leaf veins. Systemic infection and epinasty of leaves can be also observed in Nicotiana spp. Integrated PVCV sequences have been found in P. hybrida and in some wild type petunia (P. axillaris and P. integrifolia). P. parodii, on the other hand, does not contain endogenous PVCV sequences but has proved to be a suitable host to study infectivity of isolated episomal as well as chromosomal forms of PVCV (Richert-Pöggeler et al., 2003). In some cases chlorotic vein areas became necrotic during growth of infected P. parodii, which may represent a defense reaction of the new host plant following inoculation with PVCV.
- Diagnostic species
- P. hybrida gives characteristic vein-clearing symptoms (Fig. 4) 6-8 weeks after grafting of infected petunia scions or after biolistic inoculation using recombinant PVCV constructs (Richert-Pöggeler et al., 2003).
- N. tabacum (SR1) could not be infected by grafting and may therefore be a suitable non-host of episomal PVCV (Richert-Pöggeler et al., 2003).
- Propagation species
- P. hybrida cultivars that harbour multiple (and some intact) integrants of PVCV (like Himmelsroeschen or W138) are suitable propagation species, since infection can be induced from integrated viral copies (Noreen et al., 2007).
- Assay species
- No local lesion hosts have been identified so far.
Transmission by Vectors
No virus vectors have been identified to date.
Transmission through Seed
Vertical transmission of PVCV most likely occurs via viral sequences integrated into the host genome, rather than by invasion of generative tissues or seeds by episomal virus. In the progeny of infected P. parodii and N. glutinosa, which are both free of integrated PVCV sequences within their genomes, no vein-clearing symptoms developed and no PVCV specific DNA could be amplified from leaf tissue using PCR.
Transmission by Dodder
Not recorded.
Serology
A rabbit polyclonal anti-PVCV serum has been produced in spite of the limited quality (weakly immunogenic) of the purified preparations obtained (Richert, 1992). The low titered antiserum can be used to detect the virus by immunoelectron microscopy (immunosorbent electron microscopy (ISEM) and decoration) and for immunocapture PCR (IC-PCR), but is not efficient in ELISA detection (Harper et al., 2003; Richert, 1992). The same was true when monoclonal antibodies (no longer available) were tested (Richert, 1992).
Crude antiserum is useful in decoration tests at 1:50 dilutions and in immunosorbent electron microscopical trapping tests. The antiserum also contains antibodies to clathrin proteins occurring in coated vesicles of host cells. This causes problems in ELISA and Western blot applications (Richert, 1992).
No serological relations have been detected in decoration tests with antisera to CaMV, Dahlia mosaic virus (DaMV), Carnation etched ring virus (CERV) and Soybean chlorotic mottle virus (SbCMV).
Relationships
Phylogenetic analysis of reverse transcriptase domains singled out PVCV within the main cluster of the Caulimoviridae, indicating that PVCV is less related to other members of the group, than other members are to one another (Fig. 5). It is also the only known member of the family that has a genome with a single open reading frame. PVCV has therefore been classified in a distinct genus Petuvirus in the family Caulimoviridae (Fauquet et al., 2005).
Stability in Sap
Leaf sap contains few virions, which may indicate that virions are easily destroyed during the extraction procedure. Sap stability is difficult to assess since mechanical transmission is not possible.
Purification
Virus particles can be purified using the method described by Hull et al., (1976). All steps were performed at 6°C. Infected leaf tissue (100 g) was homogenised in 150 ml of 0.5 M phosphate buffer (pH 7.2) containing 0.75% sodium sulphite. After filtration, 2.5% Triton X-100 (v/v) and 1 M urea (w/v) were added and stirred for 30 minutes. Plant debris was separated out by low speed centrifugation for 10 minutes at 5000 rpm (Sorvall, GSA rotor). The supernatant was added to a 20% saccharose cushion and centrifuged for 1.5 hours at 27,000 rpm (Beckman Instruments, 45 Ti rotor). The pellet was then re-suspended in 2 ml of water overnight and purification was completed by rate zonal centrifugation in sucrose density gradients for 2 hours at 24 000 rpm (Beckman SW27 rotor). After fractionation, virion-containing portions were diluted 1:2 in 0.01 M phosphate buffer (pH 7.2), concentrated by centrifugation for 3 hours at 45,000 rpm (Beckman Ti 70 rotor) and re-suspended in 100 µl of 0.01 M phosphate buffer (pH 7.2).
Properties of Particles
Isometric particles have diameters of 43 nm when negatively stained (Fig. 1) and 46 nm in ultrathin sections (Fig. 2) (Lesemann & Casper, 1973). In comparative measurements they were significantly smaller than those of CaMV or CERV.
Particle Structure
Virions consist of isometric, non-enveloped particles with an average diameter of 43-46 nm (Fig. 1).
Particle Composition
Nucleic acid: Virions encapsidate dsDNA. As described for CaMV, different topological forms of DNA have been observed, consisting of linearised, open circular and supercoiled molecules (Richert, 1992; Richert-Pöggeler et al., 2003). The PVCV G+C content of 38% is slightly lower than the 43% G+C ratio of CaMV. As reported for CaMV, the DNA of PVCV alone is infectious. Before infection, the cloned full-length unit of viral DNA has to be released from the cloning vector. With the help of the host DNA repair machinery it is believed that these linearised molecules become circularised and can serve as templates for viral transcription and DNA synthesis. DNA biolistic inoculation has been used for transmission of PVCV (Richert-Pöggeler et al., 2003).
Coat protein: Several virus-specific bands can be detected in Western blots when hybridised with PVCV polyclonal antiserum (Richert, 1992). It is most likely that processing of PVCV structural proteins occurs, as described for CaMV. Furthermore, higher molecular bands of 90 to 95 kDa were detected that may present degradation products of clathrin heavy-chain polypeptides that have a molecular weight of 190 kDa (Robinson & Hilmer, 1990) and were co-purified during virion preparations (see also Serology).
Genome Properties
An infectious, full-length clone of episomal PVCV has been isolated from PVCV-infected N. glutinosa and sequenced (GenBank accession number U95208).
PVCV is unique among the family Caulimoviridae since all coding information is contained in one large open reading frame (Richert-Pöggeler et al., 2003). In addition to the known Caulimoviridae consensus sequences (a movement protein, an aspartic protease, an RNA binding domain, a reverse transcriptase and an RNAse H), homology to the N-terminal half of serine proteases from capilloviruses (Yoshikawa et al., 1992) was also detected in BLAST searches, in a sequence overlapping the conserved domain of a putative PVCV movement protein (Fig. 6A). The presence of multiple consensus sequences for proteases indicates post-translational processing of the polyprotein. Furthermore, Stavolone et al., (2001) identified functional coiled-coil domains, which could be part of the movement- and virion-associated proteins (Fig. 6A). Such a domain is known to be required for homo- and hetero-oligomerization and is conserved throughout the family Caulimoviridae. In CaMV, the corresponding proteins interact with each other via the coiled-coil domains, mediating virus cell-to-cell movement (Stavolone et al., 2005).
Although short stretches of similarity to sequence motifs of the catalytic domain of integrases can be identified in PVCV (Richert-Pöggeler & Shepherd, 1997), there are no further sequence homologies to recently identified putative integrase domains of retroelements (McClure et al., 2002). The 3'-untranslated region contains regulatory elements (TATA box, poly A signal) for transcription (Richert-Pöggeler & Shepherd, 1997; Noreen, 2005).
An infectious full-length copy of chromosomal PVCV (GenBank accession number AY228106, Fig. 6B) has been isolated by screening a genomic DNA library of P. hybrida (Richert-Pöggeler et al., 2003). This clone contained tandemly-arranged PVCV sequences that led to a duplication of the region upstream of the viral primer binding site, therefore representing quasi-long tandem repeats (LTRs) or QTRs. The caulimovirus QTR domain comprises promoter and polyadenylation sequences but is present only as a single copy on the circular DNA, in contrast to the pair of LTRs of retrotransposons and retroviruses. Such a head-to-tail arrangement would provide an appropriate mechanism for escape of a pararetroviral genome by recombination or, more likely, by transcription (from the promoter of the upstream copy and terminating at the polyadenylation site of the consecutive one). Systemic infection of the host plant is probably promoted by a virus-encoded movement protein. Sequence similarity to the CaMV movement protein has been identified at the N-terminus of the PVCV polyprotein (Fig. 6A).
Relations with Cells and Tissues
In ultrathin sections of symptomatic plant tissue, virions and cytoplasmic inclusion bodies can readily be detected in most of the leaf tissues except epidermal cells and sieve elements of petunia and N. glutinosa (Lesemann & Casper, 1973; Jarlfors, Richert-Pöggeler & Shepherd, unpublished). Inclusion bodies appear similar to those described for CaMV and other members of the Caulimoviridae. However, although virions of most members of the family are localised within the inclusion, those of PVCV are exclusively located in the cytoplasm surrounding the inclusion. Virus-containing regions were found to be associated with, and sometimes surrounded by, endoplasmic reticulum (Fig. 7).
Ecology and Control
Tissue culture for propagation of plant material should be avoided in Petunia breeding, since it is accompanied by wounding procedures that promote viral activation (Staginnus & Richert-Pöggeler, 2006). Furthermore, abiotic stresses like elevated temperatures or water deficiency during the growing period should be eliminated since they promote activation of endogenous PVCV copies (Noreen et al., 2007).
Recent studies indicate that chromosomal forms of PVCV are associated with DNA methylation, chromatin modification and production of small interfering (si) RNAs (Noreen et al., 2007). Therefore, RNA interference (RNAi) might be a useful tool to control integrated PVCV sequences that are potentially capable of initiating viral infection. It has been shown that dsRNA is the key molecule in triggering the plant defense system against foreign nucleic acids either by mRNA degradation and/or by methylation of DNA sequences (Mette et al., 2000; Pooggin et al., 2003). DsRNA-producing plasmids with sequence homology to the PVCV promoter region are currently under construction (Noreen, Richert-Pöggeler & Hohn, unpublished). No natural occurring resistance against PVCV is known in petunia.
Notes
Integrated PVCV sequences have been found in the pericentromeric regions of chromosomes (Richert-Pöggeler et al., 2003) and PVCV has been identified as a multicopy element within the petunia genome. The high level of sequence conservation of endogenous PVCV would suggest that PVCV integration events happened rather recently in evolutionary history. Illegitimate recombination events between host genomic DNA and viral DNA in the nucleus most likely promote multiple integrations, including tandem arrays (Salomon & Puchta, 1998). Integrated PVCV sequences are methylated (Richert-Pöggeler et al., 2003; Noreen et al., 2007) and thus may be controlled by the host.
In PCR-based detection methods, proper controls have to be included that would indicate the existence of host DNA, and thus chromosomal PVCV, as templates in the reaction (Harper et al., 2003). In general, the abundance of episomal PVCV in infected plant tissue generates a clear PCR product from crude sample preparations, whereas chromosomal copies of PVCV are under-represented and will give rise only to faint bands. Chromosomal viral sequences that have the potential to cause disease after induction have also been reported for Banana streak virus (genus Badnavirus) and Tobacco vein clearing virus (genus Cavemovirus); see review of Staginnus & Richert-Pöggeler, 2006.
Figures

Purified isometric particles of petunia vein clearing virus. Bar represents 200 nm.

Cytology of PVCV-infected leaf cells of Petunia. Main picture: overview showing three cells with dark stained cytoplasmic inclusions. Inset: enlarged view of part of an inclusion (right) and of virus particles free in the cytoplasm (left). Note absence of virus particles within the matrix of inclusion body. Magnification bars represent 1 µm and 500 nm, respectively.

Symptoms of PVCV infection in Petunia hybrida cultivar "Himmelsröschen". For comparison a leaf from a healthy plant (left) is placed next to 3 infected leaves (right) that show variation in vein-clearing symptoms as found in diseased petunia.

PVCV transmission by grafting infected scions of P. hybrida to P. parodii (Richert-Pöggeler et al., 2003). The original scion (arrow) had perished. Symptoms are epinasty of leaves, and vein-clearing. In some cases, chlorotic vein areas became necrotic during growth. Used by permission from EMBO J., 2003, 22: 4836-4845.

Phylogenetic analysis of reverse transcriptase domains of selected members of the family Caulimoviridae (Richert-Pöggeler & Shepherd, 1997). Numbers above each branch indicate the percentage of trees containing the branch in 100 bootstrap replications. Internal branches not supported by bootstrap values greater than 70 % are collapsed.
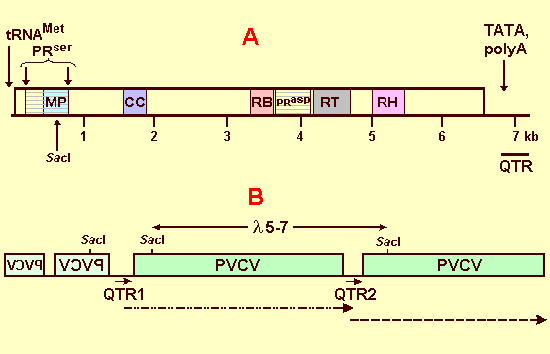
Genome organization of episomal (A) and chromosomal (B) forms of infectious PVCV sequences.
Used by permission from EMBO J., 2003, 22: 4836-4845.
A: Full-length PVCV clone (p72-2) derived from infected N. glutinosa.
Conserved domains for movement protein (MP), coiled-coil domain (CC), RNA binding domain (RB),
serine and aspartic protease (PRser and PRasp), reverse transcriptase (RT)
and ribonuclease H (RH) are indicated. Note that the PRser domain overlaps the MP domain.
CC, RB, PRasp domains are not drawn to scale. By convention, the putative tRNA binding site
for minus strand synthesis (tRNAMet) is used as the origin for nucleotide numbering.
The position of the unique SacI site is indicated. The 3'-untranslated region contains regulatory
elements (TATA box, poly A signal) for transcription. The region corresponding to the
"quasi-LTR" (QTR) is indicated.
B: Chromosomal organization of PVCV sequences in l clone 5 (17 kb)
derived from a genomic library of P. hybrida, cv. W138.
The coding regions are boxed. Flat lines represent non-coding regions within the viral genome.
PVCV sequences in reverse orientation (mirrored spelling) are found at the 5' end of the
l insert. QTRs are indicated below, SacI sites and the position of the
infectious subclone 5-7 are shown above the sequence. The interrupted lines below the QTRs indicate
position and length of putative transcripts. The 3' end of the transcript starting with QTR2 is not known,
because it was not contained within the clone. An arrow marks the length and orientation of QTR within the
untranslated region.

Cytology of PVCV-infected leaf cells of Petunia: cytoplasmic area showing freely arranged virus particles in irregular clusters and typical array of stacked elements of ER. Virus particles loosely arranged in the spaces between the ER-layers. Magnification bar represents 500 nm.
References list for DPV: Petunia vein clearing virus (417)
- Fauquet, Mayo, Maniloff, Desselberger & Ball, Virus Taxonomy: VIIIth Report of the International Committee on Taxonomy of Viruses, Elsevier Academic Press, 2005.
- Harper, Hull, Lockhart & Olszewski, Annual Review of Phytopathoogy 40: 119, 2002.
- Harper, Richert-Pöggeler, Hohn & Hull, Journal of Virological Methods, 107: 177, 2003.
- Hull, Shepherd & Harvey, Journal of General Virology 31: 93, 1976.
- Lesemann & Casper, Phytopathology 63: 1118, 1973.
- Lesemann, Petunia vein clearing virus [00.015.0.06.001.], in ICTVdB - The Universal Virus Database, version 4, ed. C. Büchen-Osmond, New York: Columbia University, 2006.
- Lockhart & Lesemann, Plant Disease 82: 262, 1997.
- McClure, Donaldson & Corro, Virology 296: 147, 2002.
- Mette, Aufsatz, van der Winden, Matzke & Matzke, EMBO Journal 19: 5194, 2000.
- Noreen, PhD thesis, Switzerland: University of Basel, 2005
- Noreen, Akbergenov, Hohn & Richert-Pöggeler, Plant Journal, 50: 219, 2007.
- Pooggin, Shivaprasad, Veluthambi & Hohn, Nature Biotechnology 21: 131, 2003.
- Richert, PhD thesis, Germany: University of Göttingen, 1992.
- Richert-Pöggeler, Noreen, Schwarzacher, Harper & Hohn, EMBO Journal 22: 4836, 2003.
- Richert-Pöggeler & Shepherd, Virology 236: 137, 1997.
- Robinson & Hillmer, Coated Pits, in The Plant Plasma Membrane, eds C. Larsson & I.M. Moller, Berlin/Heidelberg: Springer-Verlag, 1990.
- Salomon & Puchta, EMBO Journal 17, 6086, 1998.
- Staginnus & Richert-Pöggeler, Trends in Plant Science 11: 485, 2006.
- Stavolone, Herzog, Leclerc & Hohn, Journal of Virology 75: 7739, 2001.
- Stavolone, Villani, Leclerc & Hohn, Proceedings of the National Academy of Sciences USA 102: 6219, 2005.
- Yoshikawa, Sasaki, Kato & Takahashi, Virology 191: 98, 1992.
- Zeidan, Sikron, Cohen & Gera, Horticultural Science 35: 1279, 2001.