Details of DPV and References
DPV NO: 62 October 1971
Family: Rhabdoviridae
Genus: Nucleorhabdovirus
Species: Sowthistle yellow vein virus | Acronym: SYVV
Sowthistle yellow vein virus
D. Peters Department of Virology, State Agricultural University, Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described by Duffus (1963).
Synonyms
- None
A virus with enveloped, bacilliform particles about 230 x 100 nm. It has a narrow host range, is transmitted by the aphid Hyperomyzus lactucae in the persistent manner. Not transmissible by inoculation of sap. Reported only from the USA and UK.
Main Diseases
Causes vein-yellowing in Sonchus oleraceus (Fig. 1). Infection of lettuce (Lactuca sativa) has been reported (Duffus, Zink & Bardin, 1970).
Geographical Distribution
Found in the Salinas and San Joaquin Valleys in California and the Salt River Valley of Arizona. As it has also been found in England (Duffus & Russell, 1969), a wider distribution is probable.
Host Range and Symptomatology
The only known plant hosts, sowthistle and lettuce, are both members of the Compositae.
-
Diagnostic and propagation species
- Sonchus oleraceus
(sowthistle). Causes clearing of the veins or adjacent tissue (Fig. 1). - Lactuca sativa (lettuce). Vein-clearing and vein-yellowing, especially of the tips of infected leaves.
-
Assay species
- No known local lesion host. The virus has not been transmitted by inoculation of sap. Virus can be assayed by feeding the aphid vector, Hyperomyzus lactucae, on a virus suspension placed between membranes (Peters & Kitajima, 1970) or by injecting virus into the aphids (Sylvester, Richardson & Behncken, 1970). They are then tested for transmission of virus to Sonchus oleraceus plants. Primary cell cultures from Hyperomyzus lactucae were also used to assay virus suspensions (Peters & Black, 1970). Infected cells are detected by staining with fluorescent antibody.
Strains
No variants known.
Transmission by Vectors
The virus is transmitted by the aphid Hyperomyzus lactucae, in which it multiplies (Duffus, 1963; Richardson & Sylvester, 1968; Sylvester & Richardson, 1969). At 25°C the virus has a minimum incubation period in the vector of about 8 days. Transmission, except for the percentage of aphids that transmit, is independent of the length of the acquisition feeding period. The transmission efficiency is high (Duffus, 1963). The median incubation period (IP 50) at 25°C increased approximately 24 hr for every 10-fold dilution of an inoculum injected into late-instar larvae of Hyperomyzus lactucae (Sylvester et al., 1970). Transmission through the egg to some of the offspring has been reported (Sylvester, 1969).
Transmission through Seed
No seed-transmission found in Sonchus oleraceus (Peters, unpublished).
Transmission by Dodder
No information.
Serology
The virus is strongly immunogenic; rabbits immunized by three intramuscular injections of purified virus in Freund’s complete adjuvant gave antisera with titres in precipitin ring tests of 1/10,000 (Peters & Black, 1970). Other techniques were not applied by these authors.
Relationships
In particle structure, host range, mode of aphid transmission and vector specificity, the virus resembles lettuce necrotic yellows virus; however, the viruses differ in particle diameter and only lettuce necrotic yellows virus is transmissible by mechanical inoculation of sap. No serological tests for relationship between these two viruses have been reported.
Stability in Sap
The infectivity of purified virus suspensions is retained for several days at 2°C.
Purification
Peters & Kitajima (1970). Extract leaf material in 0.1 M glycine, 0.01 M MgCl2, and 0.01 M KCN at pH 8.1. Adjust the pH of the extract to 8.0 and centrifuge at 8000 g for 5 min. Add Celite to the supernatant fluid and filter the mixture through a Celite pad. Centrifuge the filtrate at 20,000 g for 1 hr. Resuspend the virus-containing pellet in 0.1 M glycine and 0.01 M MgCl2, at pH 7.0, and purify further by centrifuging the suspension at 22,500 g for 18-25 min in a density gradient (0-30% sucrose, w/v). Use zone electrophoresis in sucrose density gradients as the final step in purification. Do all steps at 2°C.
Properties of Particles
No information.
Particle Structure
Particles are bacilliform, c. 230 nm long and 100 nm wide, in preparations fixed with glutaraldehyde prior to staining with phosphotungstate or uranyl acetate (Fig. 3, Fig. 4). When fixation is omitted the particles are bullet-shaped (Fig. 2) and c. 180 nm long (Peters & Kitajima, 1970). The particles are c. 220 nm long and 95 nm wide in tissue sections (Richardson & Sylvester, 1968). The particle has an inner component consisting of a long filament coiled in a regular shallow helix (pitch 4.4 nm). The inner component seems to be bullet-shaped (Fig. 5) which may explain the disruption of the bacilliform particles into bullet-shaped particles in unfixed preparations. The envelope surface has hexagonally arranged projections. Two types of particle were distinguished: in one type the projections are in a hexagonal pattern with an angle of 30° relative to the particle axis, in the other type this angle is c. 60° (Peters & Kitajima, 1970).
Particle Composition
No information.
Relations with Cells and Tissues
In the vector, particles are found in the nucleus as well as the cytoplasm of cells of the brain, suboesophageal ganglion, salivary gland, oesophagus, ventriculus, ovaries, fat body, mycetome and muscle (Sylvester & Richardson, 1970). The particles in the nuclei are not enveloped, whereas those in the perinuclear cisternae and cytoplasm have envelopes. Masses of bacilliform particles have been found in the perinuclear spaces of infected cells of Sonchus oleraceus (Richardson & Sylvester, 1968).
Notes
The virus is readily distinguished from all others by the distinct vein-clearing produced in its hosts and by its large bacilliform particles.
Figures

(Left) infected leaf of Sonchus oleraceus, (right) healthy leaf.
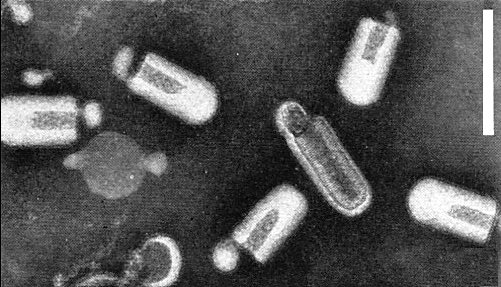
Virus particles from a purified virus preparation in neutral phosphotungstate, not previously fixed with glutaraldehyde. Bar represents 250 nm.
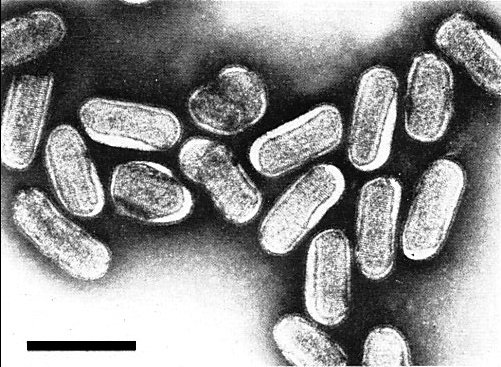
Virus particles from a purified preparation, fixed with glutaraldehyde prior to staining with neutral phosphotungstate. Bar represents 250 nm.

Particles fixed with glutaraldehyde prior to staining in uranyl acetate. Bar represents 250 nm.

Schematic representation of particle structure viewed in a plane through the long axis of the particle. The diagram shows the layer of projections, the membrane and the internal component. Bar represents 50 nm.
References list for DPV: Sowthistle yellow vein virus (62)
- Duffus, Virology 24: 194, 1963.
- Duffus & Russell, Pl. Path. 18: 144, 1969.
- Duffus, Zink & Bardin, Phytopathology 60: 1383, 1970.
- Peters & Black, Virology 40: 847, 1970.
- Peters & Kitajima, Virology 41: 135, 1970.
- Richardson & Sylvester, Virology 35: 347, 1968.
- Sylvester, Virology 38: 440, 1969.
- Sylvester & Richardson, Virology 37: 26, 1969.
- Sylvester & Richardson, Virology 42: 1023, 1970.
- Sylvester, Richardson & Behncken, Virology 40: 590, 1970.