Details of DPV and References
DPV NO: 67 October 1971
Family: Secoviridae
Genus: Waikavirus
Species: Rice tungro spherical virus | Acronym: RTSV
Rice tungro virus
G. E. Gálvez Centro Internacional de Agricultura Tropical, Apartado Aéreo 6713, Cali, Colombia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Anon. (1964) and
Rivera & Ou (1965).
Selected synonyms
-
Rice yellow orange leaf virus (Rev. appl. Mycol. 47: 169)
- Rice penyakit merah virus (Rev. appl. Mycol. 44: 3043)
- Rice leaf yellowing virus (Rev. appl. Mycol. 48: 456)
- Rice accep na pula virus (Rev. appl. Mycol. 38: 203)
- Rice mentek virus (Rev. appl. Mycol. 45: 112)
- Rice penyakit merah virus (Rev. appl. Mycol. 44: 3043)
-
A virus with isometric particles c. 30-33 nm in diameter, not transmissible mechanically but transmitted in a semi-persistent manner by the leafhoppers Nephotettix impicticeps, N. apicalis and Rocilia dorsalis. It infects a few species of Gramineae, mainly Oryza spp. It causes the most important virus diseases of rice in Southeast Asia.
Geographical Distribution
Reported from Philippines, Malaysia, Indonesia, Pakistan, Thailand and India.
Host Range and Symptomatology
Oryza spp. are the main hosts. Wathanakul (1964) found that, of 29 species of grass weeds tested, the virus produced symptoms in Eleusine indica and infected Echinochloa colonum and E. crus-galli symptomlessly. In another study (Anon., 1969), 13 of 63 graminaceous species tested were hosts of the virus but all except Oryza spp. were poor hosts of the vectors.
-
Diagnostic species
- Oryza sativa
(rice). Symptoms depend on the cultivar. The most commonly used cultivars are Taichung (Native) 1, and FK 135. Taichung (Native) 1 seedlings show stunting, with mottling and yellowing of the leaves (Fig. 1). FK 135 seedlings show conspicuous interveinal chlorosis, giving an appearance of yellow stripes with the S strain, and mottling with the M strain.Propagation species
- Taichung (Native) 1 rice plants 15-20 days after inoculation are suitable sources of virus for purification.
-
Assay species
- No local lesion host is known. One-leaf seedlings of the rice cultivar FK 135 show distinct chlorotic stripes 4-5 days after inoculation.
Strains
Two strains, severe (S) and mild (M) have been differentiated by symptoms in the rice cultivars FK 135 and Achech (Anon., 1965; Rivera & Ou, 1967).
Transmission by Vectors
The main vector is the green leafhopper, Nephotettix impicticeps (Fig. 2). The virus is also transmitted by the leafhoppers N. apicalis, N. impicticeps x N. apicalis and Recilia (Inazuma) dorsalis (Rivera & Ou, 1965; Ling, 1968, 1970; Anon., 1969, 1970). The proportion of insects that transmit the virus is about 70% for N. impicticeps, 17% for N. apicalis and 6% for R. dorsalis (Ling, 1966, 1970; Gálvez, Shikata & Miah, 1971). All five larval stages and adult leafhoppers transmit the virus in the semi-persistent manner; the proportion of transmitting insects increases with increasing acquisition access period up to 4 days (Ling, 1966). There is no latent period. The nymphs cease to transmit when they moult; all stages lose ability to transmit within 5 days after the end of an acquisition access period (50% loss after 24 h) and only regain it by renewed access to virus sources (Ling, 1966, 1969, 1970). Biotypes of N. impicticeps exist that differ in ability to infest rice cultivars (Gálvez & Miah, 1969; Anon., 1969, 1970).
Transmission through Seed
None reported.
Transmission by Dodder
No information.
Serology
No information.
Relationships
The virus is not known to be related to any other virus. The S and M strains protect against each other in plant-protection tests (Rivera & Ou, 1967).
Stability in Sap
In rice leaf extracts, the virus is stable in phosphate, phosphate-Na2 SO3, Tris, glycine, and ethylenediamine-tetraacetate (EDTA) buffers, all at 0.01 M, pH 8.0. The concentration of the virus in sap is about 1 mg/l and the thermal inactivation point (10 min) is about 60°C. The virus becomes denatured in vitro after 1 day at room temperature, 1 week at 4°C, and after 1 month when frozen. These observations on stability were made by photometric scanning of density gradient columns (Gálvez, 1968).
Purification
Homogenize infected rice leaves with 3 volumes of 0.01 M EDTA, pH 8.0. Filter and clarify the filtrate by heating at 40°C for 1 hr and centrifuging at 7000 g for 20 min. After resuspension in the same 0.01 M EDTA buffer and centrifugation at 7000 g for 20 min, the preparation may be further purified by centrifugation at 73,000 g for 3 h in sucrose density gradient columns. The virus, which forms a visible zone, may be collected and concentrated by a further cycle of differential centrifugation (Gálvez, 1968).
Properties of Particles
Sedimentation coefficient (s20,w) at infinite dilution: about 175 S. No accessory viral components are found by analytical centrifugation.
A260/A280: about 1.45 ± 0.06 (Gálvez, 1968).
Particle Structure
The particles are isometric with a diameter between 30 and 33 nm (Fig. 3).
Particle Composition
The type of nucleic acid in the particles is unknown, but if it is single-stranded the A260/A280 ratio for the purified virus suggests a nucleic acid content of about 12%; however, this seems rather little for the sedimentation coefficient (Gálvez, 1968).
Relations with Cells and Tissues
Infected cells sometimes contain dense granules and the chloroplasts and other cell components degenerate. Virus particles are found in the dense area of degenerated cells (Fig. 4) (Gálvez, Shikata & Miah, 1971).
Notes
The symptoms of rice tungro disease are similar to those of the rice transitory yellowing disease found in Taiwan, which is transmitted by the leafhoppers Nephotettix apicalis and N. cincticeps, but this virus has bacilliform particles and persists in vector leafhoppers for life.
Figures

Yellowing and stunting symptoms in naturally infected rice.
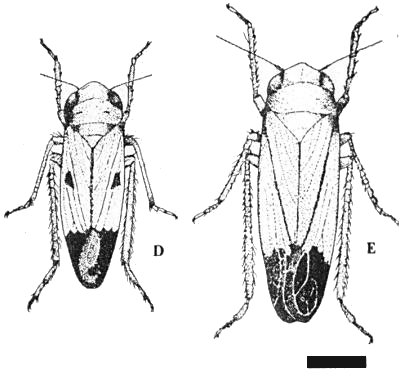
Nephotettix impicticeps, adult insects. Bar represents 1 mm. (Courtesy of International Rice Research Institute.)
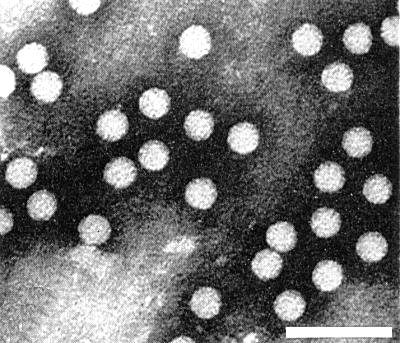
Virus particles from a purified preparation in phosphotungstate. Bar represents 100 nm.

Virus particles in an affected cell of rice. Bar represents 1 µm.
References list for DPV: Rice tungro virus (67)
- Anon., Rep. int. Rice Res. Inst. 1963: 114, 1964.
- Anon., Rep. int. Rice Res. Inst. 1964: 150, 1965.
- Anon., Rep. int. Rice Res. Inst. 1968: 103, 1969.
- Anon., Rep. int. Rice Res. Inst. 1969: 59, 1970.
- Gálvez, Virology 35: 418, 1968.
- Gálvez & Miah, Int. Rice Commn Newsl. 18: 4, 1969.
- Gálvez, Shikata & Miah, Phytopath. Z. 70: 53, 1971.
- Ling, Phytopathology 56: 1252, 1966.
- Ling, Bull. ent. Res. 58: 393, 1968.
- Ling, in Viruses, Vectors and Vegetation, New York, Inter-science Publishers, p. 255, 1969.
- Ling, J. econ. Ent. 63: 582, 1970.
- Rivera & Ou, Pl. Dis. Reptr 49: 127, 1965.
- Rivera & Ou, Pl. Dis. Reptr 51: 877, 1967.
- Wathanakul, M.S. Thesis, College Agric., Univ. Philippines, 35 pp, 1964.