Details of DPV and References
DPV NO: 74 October 1971
Family: Secoviridae
Genus: Comovirus
Species: Red clover mottle virus | Acronym: RCMV
Red clover mottle virus
V. Valenta Institute of Virology, Slovak Academy of Sciences, Bratislava, Czechoslovakia
K. Marcinka Institute of Virology, Slovak Academy of Sciences, Bratislava, Czechoslovakia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Sinha (1960).
An RNA-containing virus with isometric particles about 30 nm in diameter. It has a restricted host range and is readily transmissible by inoculation of sap; vectors are not known. It appears to be widespread, though not very common, in Europe.
Main Diseases
Causes mottle of red clover (Trifolium pratense; Fig. 2); no other natural hosts known.
Geographical Distribution
Reported from most parts of Europe except the south.
Host Range and Symptomatology
Host range is rather restricted, only a few species outside the Leguminosae being susceptible.
-
Diagnostic species
- Pisum sativum
(pea). Slight mosaic followed by chlorosis, reduction and malformation of the shoot apices. - Vicia faba (broad bean). Local necrotic lesions in inoculated leaves
followed by systemic necrosis.
- Phaseolus vulgaris (French bean). In some cultivars (e.g., some Pinto lines, Topcrop, Prince, Perlicka) local necrotic lesions (Fig. 1); in others necrotic and/or chlorotic lesions, or latent infection. Not systemic.
- Chenopodium amaranticolor and C. quinoa. Chlorotic and partially necrotic lesions. Not systemic.
- Gomphrena globosa. Necrotic lesions. Not systemic.
- Phaseolus vulgaris (French bean). In some cultivars (e.g., some Pinto lines, Topcrop, Prince, Perlicka) local necrotic lesions (Fig. 1); in others necrotic and/or chlorotic lesions, or latent infection. Not systemic.
-
Propagation species
- Pisum sativum
is a suitable plant for maintaining cultures and a good source of virus for purification.Assay species
- Phaseolus vulgaris
is a useful local lesion host.
Strains
Several minor variants, such as that of Bos & Maat (1965), can probably be distinguished.
Transmission by Vectors
No vectors known, but by analogy with the related cowpea mosaic virus, beetle vectors are likely.
Transmission through Seed
No experimental data available.
Transmission by Dodder
No experimental data available.
Serology
The virus is a good immunogen. In gel double diffusion precipitin tests, agarose (up to 1%) gives better results than agar (Valenta & Gressnerová, 1969). Several bands of precipitate may form in these tests with either rabbit hyperimmune sera or mouse immune ascitic fluid; the reason for this is not known.
Relationships
The virus is closely serologically related to several members of the cowpea mosaic virus group, especially to broad bean stain, pea green mottle, MF and cowpea mosaic viruses (Gibbs, Giussani-Belli & Smith, 1968, Valenta & Gressnerová, 1969). The relationship to other members of this group (e.g. bean pod mottle virus) is more distant.
Stability in Sap
In broad bean or French bean sap, the thermal inactivation point (10 min) is between 70 and 75°C and infectivity is retained at 20°C for more than 2 weeks (Musil & Lesková, 1969; Musil, 1966). In pea sap, the dilution end-point may be up to 10-6.
Purification
Infected pea plants may yield about 400 mg virus per kg plant tissue. Many of the purification procedures now in general use with plant viruses may be employed, but adjustment below pH 7.0 should be avoided. The following procedure gives very pure virus preparations (Marcinka, 1971):
Grind frozen tissue in 0.1 M phosphate buffer (pH 8) containing 0.02 M 2-mercaptoethanol and 10% (w/v) sucrose. Filter through gauze and clarify by low speed centrifugation. Mix supernatant with equal volume of chilled 1,1,2-trichloro- 1,2,2-trifluoro-ethane (Freon 113) and emulsify briefly but thoroughly in a blender. Separate the aqueous phase by low speed centrifugation and repeat Freon treatment twice, using one-half volumes of Freon. To the resulting aqueous phase add polyethylene glycol 6000 and NaCl to final concentrations of 6% (w/v) and 0.2 M respectively. Stir for 30 min at room temperature, sediment precipitated virus by low speed centrifugation, redissolve in 0.025 M phosphate buffer (pH 7.6) and dialyse against the same buffer for 12-24 h.
Properties of Particles
Purified preparations contain particles of uniform size which sediment as three components: RNA-free protein shells (T), which are not found with all strains of the virus, and two kinds of nucleoprotein containing different amounts of RNA (M and B). Sedimentation coefficients (s20,w) at infinite dilution (svedbergs): 60 (T), 101 (M) and 127 (B) (Gibbs, Guissani-Belli & Smith, 1968). Purified preparations of M particles are non-infective but M particles greatly enhance the infectivity of partially purified preparations of B particles (Valenta & Marcinka, 1968 and unpublished).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 13.0 (for unfractionated virus preparations).
A260/A280: 1.5 (M), 1.7 (B).
Particle Structure
Particles are isometric, about 30 nm in diameter (Fig. 3).
Particle Composition
RNA: Single-stranded; molar percentages of nucleotides in unfractionated preparations: G20.3; A29.0; C20.4; U30.2 (Gibbs, Giussani-Belli & Smith, 1968). M and B particles contain about 25 and 36% of RNA respectively (calculated from their sedimentation coefficients).
Protein: No data available.
Other components: None reported.
Relations with Cells and Tissues
Apparently all tissues are infected, including roots. Inclusion bodies are not reported and the distribution of virus within the cell is not known.
Notes
Red clover mottle virus may be confused with alfalfa mosaic virus in some hosts in which it causes similar symptoms. However, unlike red clover mottle virus, alfalfa mosaic virus is transmitted by aphids, infects Nicotiana species and has distinctive bacilliform particles.
Figures

Inoculated leaf of Phaseolos vulgaris (Perlicka), showing necrotic lesions.

Systemically infected leaf of red clover (Trifolium pratense) showing mottling, chlorotic rings and spots. Photo courtesy of Rothamsted Experimental Station.
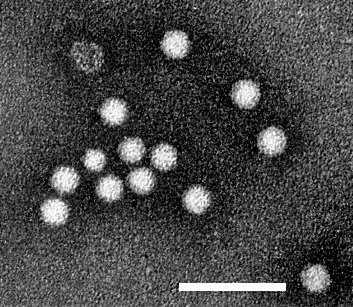
Virus particles from a purified preparation, stained with phosphotungstate. Bar represents 100 nm. Electron micrograph courtesy of Rothamsted Experimental Station.
References list for DPV: Red clover mottle virus (74)
- Bos & Maat, Neth. J. Pl. Path. 71: 8, 1965.
- Gibbs, Giussani-Belli & Smith, Ann. appl. Biol. 61: 99, 1968.
- Marcinka, Acta virol., Prague 15: 316, 1971.
- Musil, Biológia, Bratisl. 21: 663, 1966.
- Musil & Lesková Biológia, Bratisl. 24: 535, 1969.
- Sinha, Ann. appl. Biol. 48: 742, 1960.
- Valenta & Gressnerová Plant Virology, Proc. 6th Conf. Czechoslovak Plant Virologists, Academia, Prague: 62, 1969.
- Valenta & Marcinka, Acta virol., Prague 12: 288, 1968.