Details of DPV and References
DPV NO: 77 October 1971
Family: Virgaviridae
Genus: Furovirus
Species: Soil-borne wheat mosaic virus | Acronym: SBWMV
Soil-borne wheat mosaic virus
M. K. Brakke Agricultural Research Service, US Department of Agriculture, Plant Pathology Department, University of Nebraska, Lincoln, Nebraska 68503 USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
McKinney (1923).
Selected synonyms
- Marmor tritici
(McKinney, 1944) - Wheat viruses 1 and 3 (McKinney, 1937)
-
An RNA-containing virus with stiff rod-shaped particles of two lengths, 110-160 and 300 nm, which appear hollow when negatively stained. It has a narrow host range, is transmitted by the fungus, Polymyxa graminis, and by inoculation of sap and causes mosaic diseases in winter wheat and barley.
Main Diseases
Causes mosaic diseases in winter wheat (Fig. 3) and barley, the severity of symptoms depending on variety, strain of virus, and weather. Mosaic may be light green to yellow. Roots are usually stunted. Some varieties of wheat, e.g. those related to Harvest Queen, show rosetting, with extreme stunting of tops and roots and excessive tillering. Symptoms normally appear on new growth of winter wheat in the spring, but have been seen in occasional years in the autumn. However, virus can be detected in the autumn in the roots of young plants not showing leaf symptoms (Brakke, Estes & Schuster, 1965). Affected areas of the field appear yellow or light green from a distance. The disease tends to be more severe in low-lying areas of a field, but is not confined to these areas. Symptoms become milder and tend to disappear as the weather becomes warmer and the wheat plants larger (Koehler, Bever & Bonnett, 1952).
Geographical Distribution
Winter wheat-growing areas of USA, Japan and Italy.
Host Range and Symptomatology
The host range of US strains is limited to several Triticum (wheat) species, Hordeum vulgare (barley), Secale cereale (rye), Bromus commutatus, B. tectorum and some Chenopodium species (McKinney, 1944; Rao & Brakke, 1969; Paulsen, 1970). Agropyron repens, Bromus inermis, Avena sativa (oats), A. byzantina, Zea mays (maize), Nicotiana tabacum, Lycopersicon esculentum, Cucumis sativus, and Phaseolus vulgaris are not hosts (McKinney, 1944).
-
Diagnostic species
- Triticum aestivum
(wheat). The virus is manually transmissible to wheat, producing symptoms in 2 weeks or more at 20°C or less, but few or no symptoms above 20°C. The optimum temperature for symptoms in the greenhouse, or in growth chambers, is about 16°C, with an optimum day length of about 8 hr. Infected plants show a green or yellow mosaic and stunting. It is difficult to infect all the inoculated plants by leaf-rubbing. Inocula prepared by grinding infected leaves in 0.1 M K2HPO4 are usually more infectious than those prepared by grinding leaves in water. Air-brush inoculation gives a higher percentage infection than leaf-rubbing (Pring & Gumpf, 1970). The virus is translocated slowly in the plant. Seedlings with two leaves or more, inoculated either by leaf-rubbing, or through the roots by the vector, seldom show mosaic symptoms; the virus apparently remains localized in roots or crown. The percentage of inoculated plants showing symptoms is increased when the plants are placed in the dark for 4-5 days starting a few days after inoculation, or when the leaves are cut off after a month and new growth observed (Rao & Brakke, 1970).Propagation species
- Wheat (cv. Michigan Amber and many others).
-
Assay species
- Systemic assays on wheat (cv. Michigan Amber or others) and local lesions on Chenopodium quinoa and C. amaranticolor.
Strains
Green and yellow mosaic strains and a rosetting strain were selected by McKinney (1931). The degree of relationship between apparently similar viruses from the USA and Italy has not been determined. Isolates from Japan (the soil-borne green mosaic of wheat and barley) and USA are serologically related but those from Japan can infect tobacco and Zea mays (Tsuchizaki, Hibino & Saito, 1970).
Transmission by Vectors
McKinney, Paden & Koehler (1957) suggested that a vector transmitted this virus in soils because (i) some chemical treatments eradicated the virus from soils, and (ii) steamed soil in which manually inoculated wheat plants grew did not become infective whereas soil mixed with washed, naturally infected roots did (McKinney, 1948). Similar results were reported by Saito et al. (1964c, 1964d). Linford & McKinney (1954) suggested that the fungus, Polymyxa graminis (Fig. 2, Fig. 4), found in roots of plants infected through the soil, might be a vector. Estes & Brakke (1966) established correlations between transmission of the virus and the presence of P. graminis. Canova (1966) reported that cultures of P. graminis transmitted the soil-borne wheat mosaic virus in Italy. Virus transmission was not prevented by treating zoospores of P. graminis with antiserum to the virus, nor by treating resting spores of the fungus with 0.1 N NaOH or 0.1 N HCl. Zoospores could not be freed of virus by washing by repeated centrifugation. Zoospores did not acquire virus in vitro, but virus-free P. graminis acquired virus when grown in plants infected with virus by manual inoculation (Rao & Brakke, 1969). The infectivity in soil survived air-drying (McKinney, 1937).
Transmission through Seed
Not seed-transmitted.
Serology
The virus is very immunogenic in rabbits; injection of purified virus results in antisera with precipitation end-points of 1/1000 or more. The end-point in precipitin reactions depends on the virus preparation, being higher in preparations where the virus has partly aggregated before addition of antisera. Reactions have also been obtained in agar diffusion tests (Saito, Takanashi & Iwata, 1964b; Gumpf, 1970; Rao & Brakke, 1969).
Relationships
The virus is morphologically similar to other tubular viruses, in particular tobacco mosaic, tobacco rattle, and barley stripe mosaic viruses. It appears to be unrelated to some other soil-borne viruses in cereals, such as wheat spindle streak mosaic (Slykhuis, 1970), the soil-borne oat mosaic viruses occurring in USA and UK, and to the yellow mosaic viruses of barley and wheat in Japan (Inouye, 1969), all of which reportedly have flexuous filamentous particles.
Stability in Sap
The thermal inactivation point in wheat sap is 60-65°C (10 min) and dilution end-point is 10-2-10-3. The virus survived more than 11 years in dried leaves (McKinney, Silber & Greeley, 1965).
Purification
The main difficulty in purifying the virus is its tendency to aggregate. It has been purified by differential centrifugation after clarifying the plant extracts with chloroform (Saito et al., 1964a), or chloroform and ethylene dichloride (Gumpf, 1971). It has also been purified by differential centrifugation followed by treatment with detergent to disperse chloroplast fragments and other impurities (Rao & Brakke, 1969). The virus appears to be extracted better with 0.5 M sodium orthoborate, pH 9.0, than with extractants of lower pH and salt concentration. The yield is up to 150 mg per kg of tissue (Gumpf, 1970).
Properties of Particles
The following properties were determined for the Nebraska isolate (Gumpf, 1971).
Sedimentation coefficient, (s20,w): 172 S (160 nm rods); 211 S (300 nm rods).
A260 for 1 mg/ml, 1 cm light path: 3.1 (not corrected for light scattering).
A260/A280: 1.20 (not corrected for light scattering).
Particle Structure
Particles are stiff rods (Fig. 1), appearing hollow, like those of tobacco mosaic and tobacco rattle viruses, when negatively stained. The diameter is about 20 nm. The most frequent length is 150-160 nm (Tsuchizaki et al. (1970) reported 110-120 nm for some Japanese isolates), but many are about 300 nm long (Brakke et al., 1965; Brandes, Phillipe & Thornberry, 1964). Some of the particles in preparations of the 300 nm rods appear to be dimers of the 160 nm rods. Such preparations are infective, but preparations of 160 nm rods are not (Gumpf, 1971).
Particle Composition
Nucleic acid: The Nebraska isolate contains two classes of single-stranded RNA, with sedimentation coefficients (s20,w) of 24.3 and 33.4 S in 0.15 M NaCl, 0.015 M sodium citrate, pH 7.0. Their respective M. Wts are 1.0 x 106 and 2.0 x 106, based on their sedimentation rate after formaldehyde treatment. The 160 nm rods contain only the 24.3 S RNA. Preparations of the 300 nm rods have both classes of RNA, the 24.3 S RNA coming from dimers of 160 nm rods and the 33.4 S RNA from true 300 nm rods. There is always more of the 24.3 S RNA than of the 33.4 S, but the ratio is variable. After one centrifugation through sucrose gradients, which may be insufficient for complete separation, the 33.4 S RNA is infective, hut the 23.4 S RNA is not (Gumpf, 1971). The A260/A280 ratio suggests that the particles have about 5% RNA.
Protein: No information.
Relations with Cells and Tissues
The virus infects both roots and leaves. McKinney, Eckerson & Webb (1923) observed vesicular, amorphous inclusions by light microscopy, whereas three types of inclusion have been observed by electron microscopy. Crystalline inclusions, apparently composed of virus particles, were frequently found in the vacuole. The other two types of inclusion were found in the cytoplasm. One of these was apparently crystalline and had a smaller lattice spacing than would be expected if it contained virus; it was surrounded by virus particles. The other type of inclusion was amorphous and contained convoluted tubules and scattered virus particles (Peterson, 1970).
Notes
The particles of this virus vary in length with the strain (Tsuchizaki et al., 1970), as happens with tobacco rattle virus. The wide range of lengths reported for ‘soil-borne wheat mosaic virus’ (see Brandes et al., 1964, for references) could also indicate this, but it is not certain that all these reports refer to strains of the same virus. Lengths of rods, and the size and function of the RNA components of different strains should be compared. Perhaps the best way to recognize this virus is by symptoms, together with the pattern of infected areas in the field, and the numerous rods found by electron microscopy of leaf-dip preparations. Barley stripe mosaic virus is the only other virus with a similar morphology found in wheat; it differs in having a shorter incubation period after manual transmission to wheat and in giving good symptoms at 20-25°C.
Figures

Soil-borne wheat mosaic virus in leaf-dip preparation mounted in a 3:1 mixture of 2% potassium phosphotungstate: 1% ammonium vanadatomolybdate. Bar represents 200 nm.
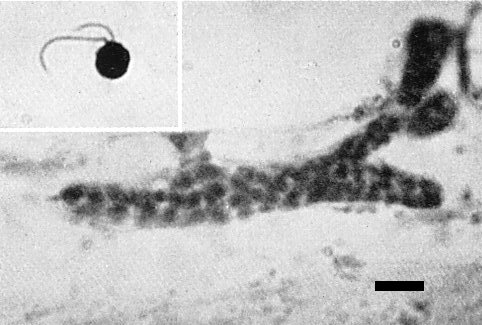
Polymyxa graminis sporangium with exit tube reaching host cell wall. Bar represents 12 µm. Inset is a biflagellate zoospore of P. graminis at 2.5 x the magnification of the sporangium. (From Rao, 1968.)
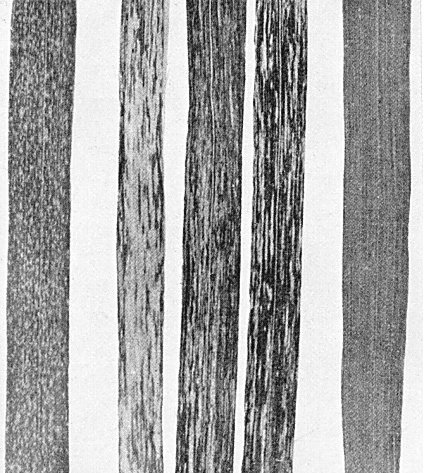
Wheat leaves; (left) a leaf showing non-parasitic mottling; (centre) three leaves showing soil-borne mosaic; (right) a healthy leaf. (From Koehler et al., 1952.)
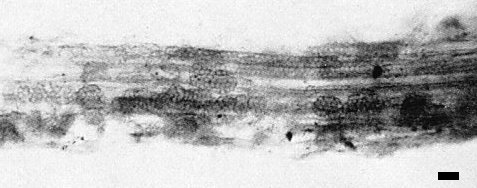
P. graminis resting spores in a wheat root, unstained. Bar represents 20 µm. (Photograph by Jean Peterson.)
References list for DPV: Soil-borne wheat mosaic virus (77)
- Brakke, Estes & Schuster, Phytopathology 55: 79, 1965.
- Brandes, Phillipe & Thornberry, Phytopath. Z. 50: 181, 1964.
- Canova, Phytopath. Mediterranea 5: 53, 1966.
- Estes & Brakke, Virology 28: 772, 1966.
- Gumpf, Ph.D. Thesis, University of Nebraska, Lincoln, 1970.
- Gumpf, Virology 43: 588, 1971.
- Inouye, Ber. Ohara Inst. landw. Biol. 15: A7, 1969.
- Koehler, Bever & Bonnett, Bull. Ill. agric. Exp. Stn 556: 567, 1952.
- Linford & McKinney, Pl. Dis. Reptr 38: 711, 1954.
- McKinney, J. agric. Res. 23: 771, 1923.
- McKinney, Science, N.Y. 73: 650, 1931.
- McKinney, Circ. U.S. Dep. Agric. 442, 22 pp., 1937.
- McKinney, J. Wash. Acad. Sci, 34: 322, 1944.
- McKinney, Phytopathology 38: 1003, 1948.
- McKinney, Eckerson & Webb, J. agric. Res. 26: 605, 1923.
- McKinney, Paden & Koehler, Pl. Dis. Reptr 41: 256, 1957.
- McKinney, Silber & Greeley, Phytopathology 55: 1043, 1965.
- Paulsen, Phytopathology 60: 1307, 1970.
- Peterson, Virology 42: 304, 1970.
- Pring & Gumpf, Pl. Dis. Reptr 54: 550, 1970.
- Rao, Phytopathology 58: 1516, 1968.
- Rao & Brakke, Phytopathology 59: 581, 1969.
- Rao & Brakke, Phytopathology 60: 714, 1970.
- Saito, Takanashi, Iwata & Okamoto, Bull. natn. Inst. agric. Sci., Tokyo Ser. C 17: 1, 1964a.
- Saito, Takanashi & Iwata, Bull. natn. Inst. agric. Sci., Tokyo Ser. C 17: 23, 1964b.
- Saito, Takanashi, Iwata & Okamoto, Bull. natn. Inst. agric. Sci., Tokyo Ser. C 17: 41, 1964c.
- Saito, Takanashi, Iwata & Okamoto, Bull. natn. Inst. agric. Sci., Tokyo Ser. C 17: 61, 1964d.
- Slykhuis, Phytopathology 60: 319, 1970.
- Tsuchizaki, Hibino & Saito, Ann. phytopath. Soc. Japan 36 : 187, 1970.