Details of DPV and References
DPV NO: 92 June 1972
Family: Bromoviridae
Genus: Cucumovirus
Species: Peanut stunt virus | Acronym: PSV
Peanut stunt virus
G. I. Mink Washington State University, Irrigated Agriculture Research and Extension Center, Prosser, Washington 99350, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Troutman (1966a,
1966b) and
Silbernagel, Mink & Saksena (1966).
Synonym
- Groundnut stunt virus (Rev. appl. Mycol. 47: 2420)
-
An RNA-containing virus with isometric particles about 30 nm in diameter. It has a wide host range, is transmitted by aphids in the non-persistent manner and is readily transmissible by inoculation of sap. Found in USA and Japan.
Main Diseases
Causes pronounced stunting of peanuts (Arachis hypogaea) and malformation of the fruit (Troutman, 1966a); epinasty, leaf distortion and stunt of beans (Echandi & Hebert, 1971; Mink, Silbernagel & Saksena, 1969); chlorotic mottle and stunt of Burley tobacco (Gooding, 1968) and systemic mottle of several clovers (Choopanya & Halpin, 1968). Naturally infected white clover (Trifolium repens) may be the overwintering source of the virus in North Carolina (Hebert, 1967).
Geographical Distribution
Economically important in North Carolina and Virginia, USA. Also reported to occur in the states of Georgia (Kuhn, 1971) and Washington (Silbernagel et al., 1966), and in Japan (T. Tsuchizaki, unpublished).
Host Range and Symptomatology
Transmissible by inoculation of sap to many dicotyledonous species. Many leguminous plants are susceptible, including all varieties of Phaseolus vulgaris tested (Zaumeyer & Goth, 1967). One or more species of Chenopodiaceae, Compositae, Cucurbitaceae and Solanaceae are also susceptible.
-
Diagnostic species
- Vigna sinensis
(cowpea). Chlorotic lesions on primary leaves 4-5 days after inoculation (Fig. 3); severe epinasty and vein-clearing of trifoliolate leaves 2-3 days later. - Phaseolus vulgaris (French bean). Symptoms depend on the variety; lesions on inoculated
primary leaves may be chlorotic or necrotic; systemically infected trifoliolate leaves develop
chlorotic mottle or mosaic. The variety Bountiful is a useful indicator, producing elongated and
mis-shapen trifoliolate leaves (Fig. 1).
- Datura stramonium (jimson weed). Systemic chlorotic mottle with islands of green tissue (Fig. 4).
- Lycopersicon esculentum (tomato). Strap-like malformation of leaflets producing a fernleaf effect similar to that caused by infection with cucumber mosaic or tomato aspermy viruses (Fig. 2).
- Chenopodium amaranticolor and C. quinoa. Chlorotic local lesions with no systemic invasion.
- Nicotiana tabacum (tobacco). Light green to yellow rings 5-10 mm in diameter with broad margins on the inoculated leaves followed by chlorotic areas in the younger leaves.
- Datura stramonium (jimson weed). Systemic chlorotic mottle with islands of green tissue (Fig. 4).
-
Propagation species
- Vigna sinensis is useful both for maintaining cultures and as a source of virus for purification.
-
Assay species
- Vigna sinensis, Phaseolus vulgaris, Chenopodium amaranticolor
and C. quinoa are useful local lesion hosts.
Strains
Minor variants, differing in one or more host reactions, can be distinguished. Two major strains have been reported (Mink, Hebert & Silbernagel, 1967) which differ in host range, serological relationships and particle stability:
Eastern strain. Found in southeastern USA. Produces necrotic primary lesions, systemic necrotic rings and veinal necrosis in Tennessee Greenpod bean, and mosaic without necrosis in Perfected Wales pea.
Western strain. Found in Washington State, USA. Produces mosaic without necrosis in Tennessee Greenpod bean, and mosaic with severe necrosis of stems and petioles in Perfected Wales pea.
Transmission by Vectors
Transmitted in the non-persistent manner by the aphids Aphis craccivora, A. spiraecola and Myzus persicae but not by A. gossypii (Hebert, 1967). Virus can be acquired in less than 1 min and inoculated in less than 1 min. No latent period. Not transmitted to progeny aphids. Some isolates lose their aphid-transmissibility after serial transfer by inoculation of sap; one such isolate apparently regained aphid-transmissibility (S. Tolin, unpublished).
Transmission through Seed
Transmitted through 0.1% or less of the seed from infected peanut plants (Troutman, Bailey & Thomas, 1967; Kuhn, 1969). Because seedlings from infected seed emerge late and grow poorly, seed transmission is not considered an important factor in disease spread.
Transmission by Dodder
Transmitted by an unidentified species of dodder (Miller & Troutman, 1966).
Serology
The virus is not strongly immunogenic in rabbits. Ionic conditions appear to affect particle stability before and after injection, but this has not been adequately studied. Both ring interface and gel-diffusion tests can be used. Virus in crude sap can be detected by gel-diffusion tests; the Eastern strain produces a single curved band near the antigen well, whereas the Western strain produces a curved band near the antigen well and one or more straight bands nearer the antiserum well. The number, intensity and location of bands produced in gel-diffusion plates by purified virus, especially the Western strain, is dependent upon the previous treatment of the sample and on the antiserum used (Mink et al., 1969).
Relationships
Many isolates from southeastern USA are serologically identical (Echandi & Hebert, 1971; Gooding, 1968; Kuhn, 1969), but one differs slightly serologically from the others (S. A. Tolin, unpublished). Isolates from Washington State differ greatly from those from southeastern USA (Mink et al., 1967).
The physical and chemical properties of peanut stunt virus suggest affinities with cucumber mosaic or tomato aspermy viruses, and peanut stunt virus is in fact related serologically to some strains of both viruses. However, strains of all three viruses differ serologically among themselves, and the degree of apparent relationship therefore depends upon the particular strain of each virus and the particular antiserum used. In gel-diffusion tests, both strains of peanut stunt virus react with some antisera to tomato aspermy virus, giving reactions of partial identity. However, only one of these antisera, prepared against the isolate of Blencowe & Caldwell (1949), gave reactions suggesting a direct serological relationship between peanut stunt virus and cucumber mosaic virus. Some antisera to cucumber mosaic virus possess type-specific antibodies to one or both strains of peanut stunt virus, but no antibodies that react both with the homologous virus and with peanut stunt virus (Mink, 1969).
Antiserum against the Western strain of peanut stunt virus does not react with cucumber mosaic virus antigens. Antiserum against the Eastern strain reacted only with a low molecular weight antigen associated with the cowpea strain of cucumber mosaic virus in gel-diffusion plates (Mink, 1969); however, J. W. Groelke & S. A. Tolin (pers. comm.) obtained reciprocal heterologous reactions between the Eastern strain of peanut stunt virus and cucumber mosaic virus using a modification of the test described by Ball & Brakke (1969) in which antigen-antibody reaction products were detected by centrifugation in sucrose density gradients.
Stability in Sap
Infectivity is lost after 4-24 h at room temperature depending upon the source plant used. Addition of antioxidants or reducing agents preserves infectivity for several days. Thermal inactivation occurs after 10 min at 50-60°C, the exact temperature depending upon the strain tested. Dilution end-point is between 10-3 and 10-4.
Purification
Harvest primary leaves of Vigna sinensis 7-8 days after inoculation. Purify by Steere’s chloroform-butanol method followed by differential ultracentrifugation. An unidentified yellow pigment is removed by rate zonal sucrose density-gradient centrifugation. Yield and specific infectivity are very dependent on the virus strain, the clarification procedure and the ionic conditions of the suspending medium (Mink et al., 1969; Tolin, 1967, 1969).
Properties of Particles
Sedimentation coefficient, s20, w: about 98 S.
Molecular weight: about 6.8 x 106 daltons.
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 4.8.
A260/A280: 1.64 before dialysis, 1.58 after dialysis against cold 0.01 M phosphate buffer at pH 8.0.
Particle Structure
The particles are isometric, about 30 nm in diameter (Fig. 5). The particles are penetrated by phosphotungstate and contrast poorly.
Particle Composition
RNA: Molecular weight about 1 x 106, about 16% of the particle weight, probably single-stranded. Molar percentages of nucleotides: G24, A26, C21, U29. Sedimentation coefficient of RNA is about 23 S when calculated with reference to whole virus in sucrose density-gradients containing 0.01 M phosphate buffer, pH 8.0. Infectious RNA is readily obtained from the Western strain but not from the Eastern strain by freezing purified virus in 2 M LiCl.
Protein: Amino acid composition (moles %): ala 7.2; arg 6.6; asx 8.7; cys 1.0; glu 7.9; gly 5.7; his 3.0; ile 3.3; leu 8.4; lys 6.6; met 0; phe 3.0; pro 7.4; ser 10.2; thr 8.2; trp 7.0; tyr 3.0; val 9.3 (J. H. Tremaine, unpublished).
Other components: None reported.
Relations with Cells and Tissues
No information.
Notes
In physical and chemical properties peanut stunt virus clearly belongs to the same group as cucumber mosaic virus, tomato aspermy virus, the aspermy-related viruses from chrysanthemum and, possibly, robinia mosaic virus, although the particle size of robinia mosaic virus seems larger than that of other members of this group (Schmelzer, 1967). The characteristics of strains and viruses in this group overlap to such an extent that no single property is sufficiently reliable for identification. Peanut stunt virus strains show various degrees of serological relationship with strains of tomato aspermy virus and cucumber mosaic virus but they do not fit the scheme proposed by Lawson (1967) to distinguish aspermy viruses from cucumber mosaic virus strains. Unlike tomato aspermy virus, peanut stunt virus strains systemically infect cucumber, French bean and Datura stramonium and do not cause enations on leaves of Nicotiana species. Peanut stunt virus can be differentiated from cucumber mosaic virus by the systemic reactions it produces in cowpea, French bean and peanut, and from robinia mosaic virus by systemic infection of Petunia hybrida. Serological tests, particularly gel-diffusion tests, may give confusing or misleading results on inter-relationships in this group due to qualitative differences among different antisera prepared to a given virus and to the presence of antibodies apparently specific to peanut stunt virus, but not to tomato aspermy virus, in many antisera to cucumber mosaic virus (Mink, 1969).
Figures

Systemically infected trifoliolate leaf of Bountiful bean.

Systemically infected leaf of Bonny Best tomato.

Chlorotic lesions on inoculated primary leaves of cowpea.

Systemically infected leaf of Datura stramonium.
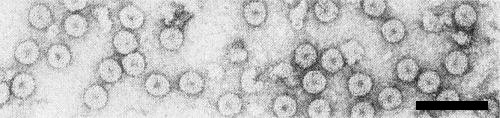
Virus particles from a purified preparation stained with 1% uranyl acetate. Bar represents 100 nm. (Courtesy S. A. Tolin.)
References list for DPV: Peanut stunt virus (92)
- Ball & Brakke, Virology 39: 746, 1969.
- Blencowe & Caldwell, Ann. appl. Biol. 36: 320, 1949.
- Choopanya & Halpin, Phytopathology 58: 726, 1968.
- Echandi & Hebert, Phytopathology 61: 328, 1971.
- Gooding, Phytopathology 58: 728, 1968.
- Hebert, Phytopathology 57: 461, 1967.
- Kuhn, Phytopathology 59: 1513, 1969.
- Kuhn, Pl. Dis. Reptr 55: 453, 1971.
- Lawson, Virology 32: 357, 1967.
- Miller & Troutman, Pl. Dis. Reptr 50: 139, 1966.
- Mink, Phytopathology 59: 1889, 1969.
- Mink, Hebert & Silbernagel, Phytopathology 57: 1400, 1967.
- Mink, Silbernagel & Saksena, Phytopathology 59: 1625, 1969.
- Schmelzer, Phytopath. Z. 58: 59, 1967.
- Silbernagel, Mink & Saksena, Phytopathalogy 56: 901, 1966.
- Tolin, Phytopathology 57: 834, 1967.
- Tolin, Phytopathology 59: 1560, 1969.
- Troutman, Phytopathology 56: 587, 1966a.
- Troutman, Phytopathology 56: 904, 1966b.
- Troutman, Bailey & Thomas, Phytopathology 57: 1280, 1967.
- Zaumeyer & Goth, Phytopathology 57: 837, 1967.