Details of DPV and References
DPV NO: 95 June 1972
Family: Potyviridae
Genus: Potyvirus
Species: Henbane mosaic virus | Acronym: HMV
Henbane mosaic virus
D. A. Govier Rothamsted Experimental Station, Harpenden, Hertfordshire, England
R. T. Plumb Rothamsted Experimental Station, Harpenden, Hertfordshire, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Hamilton (1932) and Lovisolo & Bartels (1970).
- Selected synonyms
- Hyoscyamus virus III (Rev. appl. Mycol. 17: 64)
- Hyoscyamus virus I (Rev. appl. Mycol. 17: 52)
- Atropa mild mosaic virus (Rev. Pl. Path. 50: 2197)
- Hyoscyamus virus I (Rev. appl. Mycol. 17: 52)
An RNA-containing virus with filamentous particles, 800 or 900 nm long (see below) that infects mainly solanaceous hosts. It is readily transmitted by inoculation of sap, and by aphids in the non-persistent manner.
Main Diseases
Causes mosaic diseases in Hyoscyamus niger, Datura spp. and Physalis alkekengi, and strains have been isolated from Atropa belladonna.
Geographical Distribution
Reported only from England, Germany and Italy although it may be widespread in solanaceous weed hosts. Not reported naturally infecting tobacco.
Host Range and Symptomatology
Host range mainly in the Solanaceae. Readily transmitted by inoculation of sap to:
- Diagnostic species
- Hyoscyamus niger
(Fig. 1, Fig. 2), Nicotiana tabacum (Fig. 3), N. glutinosa and N. rustica. Inoculated leaves usually develop poorly defined chlorotic or necrotic lesions. Vein-clearing of the young leaves is followed by a green or yellow-green systemic mosaic, often with dark green blistering and sometimes with necrotic spots or rings. - Nicotiana sylvestris. Most strains give necrotic lesions, often in
the form of rings, on the inoculated leaves (Fig. 5); systemic mottling and
necrosis develop later.
- Datura stramonium. Severe systemic mosaic with leaf-narrowing, blistering and malformations (Fig. 6).
- Chenopodium amaranticolor. Some strains give circular chlorotic local lesions with necrotic centres; not systemic.
- Solanum demissum x S. tuberosum ‘A-6’. Necrotic local lesions (Fig. 4).
- Datura stramonium. Severe systemic mosaic with leaf-narrowing, blistering and malformations (Fig. 6).
- Propagation species
- Nicotiana tabacum
cv. Xanthi-nc may be used for maintaining the virus and is a good source of virus for purification.- Assay species
- S. demissum
x S. tuberosum ‘A-6’, N. sylvestris.
Strains
Many minor variants have been obtained from the original isolate (Watson, 1968) and these and the strains isolated from naturally infected Atropa belladonna (Bode, Brandes & Paul, 1969; Harrison & Roberts, 1971; Govier & Woods, 1971), Datura spp. (Bradley, 1952; Lovisolo & Bartels, 1970) and Physalis alkekengi (Lovisolo & Bartels, 1970) differ from the type virus in their reactions on some hosts.
Transmission by Vectors
Most strains are transmitted readily by Myzus persicae; M. ascalonicus, Aulacorthum circumflexum and Macrosiphum euphorbiae are also vectors (Watson & Roberts, 1939; Doncaster & Kassanis, 1946). All instars can transmit. Aphids can acquire the virus in 10 sec and inoculate it in a similar period. There is no latent period. Some feeding aphids retain infectivity for 6 hr and starving aphids for 24 hr. Not transmitted to progeny aphids.
Transmission through Seed
Not reported.
Transmission by Dodder
Not reported.
Serology
Strongly immunogenic. The virus gives flocculent precipitates in tube-precipitation tests and is readily detected in tobacco sap by slide-agglutination tests. Agar gel-diffusion tests fail with intact virus.
Relationships
Strains occurring naturally in Atropa belladonna, Datura spp. and Physalis alkekengi are closely related serologically to isolates derived from the original henbane culture. In plant-protection tests, strains protect against one another. Discrepancies in particle length determinations, and the possibility that some cultures of henbane mosaic virus were contaminated with potato virus Y, cast some doubt (Govier & Woods, 1971) on reports that henbane mosaic virus is distantly related serologically to several viruses of the potato virus Y group (Bartels, 1964), but the serological relationship with potato virus Y has now been confirmed (D. A. Govier, unpublished). Distant serological relationships have also been reported with Columbian datura virus (Kahn & Bartels, 1968) and pokeweed mosaic virus (Shepherd, Fulton & Wakeman, 1969).
Stability in Sap
In tobacco or Datura stramonium sap, the thermal inactivation point (10 min) is about 60°C, dilution end-point about 10-6, and most infectivity is lost in a few days at 20°C.
Purification
The following method (D. A. Govier & B. Kassanis, unpublished) gives infective preparations in which the particles are less aggregated than in preparations made by other methods. Homogenize leaves in a cooled blender with 0.1 M ethylenediamine-tetraacetate pH 7.6 containing 0.01 M Na-diethyldithiocarbamate (DIECA), 2 ml/g tissue. Squeeze the sap through muslin, clarify by centrifuging for 30 min at 15,000 g, dialyse for 18-24 hr against 10 vol of 0.005 M DIECA and reclarify. Shake the supernatant fluid for 5-10 min with 1/4 vol of chloroform, centrifuge, and subject the aqueous phase to 2 or 3 cycles of differential centrifugation (50 min at 100,000 g, 10 min at 15,000 g), resuspending virus pellets in 0.01 M borate pH 7.5. Aggregation is largely prevented by leaving pellets covered with buffer overnight at 5°C before resuspending, by concentrating only slightly when resuspending the first high-speed pellets and, when a large weight of leaves is used, by infiltrating the leaves with extracting solution under vacuum before homogenizing them. About 30 mg of virus are obtained from 1 kg of leaves.
Properties of Particles
Sedimentation coefficient (s20,w) at infinite dilution: about 160 S.
A260/A280: 1.1.
Particle Structure
Particles are filamentous, 12-13 nm wide and helically constructed, their length and appearance depending on the conditions of extraction (Govier & Woods, 1971). When magnesium ions are absent, the particles are flexuous and have a normal length of about 800 nm (Fig. 7), but when magnesium is present, the particles are straight or only slightly flexuous and have a normal length of about 900 nm (Fig. 8).
Particle Composition
RNA: Molar percentages of nucleotides G26; A29; C27; U18 (J. M. Carpenter, unpublished).
Protein: Subunits have molecular weight of about 3.2 x 104 (J. M. Carpenter, unpublished).
Relations with Cells and Tissues
Details reported for the atropa mild mosaic strain (Harrison & Roberts, 1971) have been confirmed for the type strain (R. T. Plumb & D. A. Vince, unpublished). Pinwheel inclusions occur in sections of systemically infected tissue and in leaf-dip preparations, often with virus particles lying parallel to their long axis. The pinwheel lamellae have characteristic striations with a spacing of about 5 nm. Crystalline material with a honeycomb-like structure occurs in sections of infected cells. Virus-like particles show in leaf sections fixed in glutaraldehyde and osmic acid (Harrison & Roberts, 1971), or better in sections fixed in osmic acid alone (Plumb & Vince, 1971).
Notes
General similarity of properties, the presence of pinwheel inclusions in infected cells and distant serological relationships, place this virus in the potato virus Y group, although its particles are longer than those of other viruses in the group. In contrast to potato virus Y, it infects Datura stramonium. It is readily distinguished from tobacco etch virus because it does not produce intranuclear inclusions.
Figures

Diseased plant of Hyoscyamus niger.

Systemically infected leaf of Hyoscyamus niger.

Systemically infected leaf of Nicotiana tabacum cv. Xanthi-nc.

Inoculated leaf of Solanum demissum x S. tuberosum A-6, showing local lesions. (Courtesy R. Bartels.)

Inoculated leaf of Nicotiana sylvestris, showing local lesions.
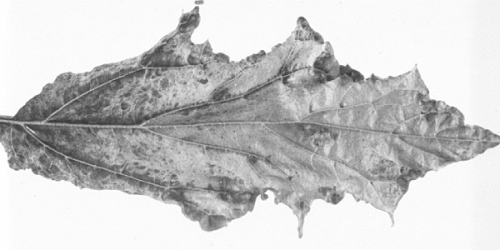
Systemically infected leaf of Datura stramonium.

Virus particles from a purified preparation stained with phosphotungstate. Bar represents 100 nm.
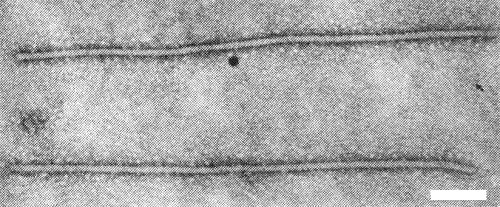
Virus particles from the same preparation after adding Mg ions. Stained with phosphotungstate. Bar represents 100 nm.
References list for DPV: Henbane mosaic virus (95)
- Bartels, Phytopath. Z. 49: 257, 1964.
- Bode, Brandes & Paul, Jber. biol. BundAnst. Land- u.Forstw. Braunschweig 1968: A61, 1969.
- Bradley, Ann. appl. Biol. 39: 78, 1952.
- Doncaster & Kassanis, Ann. appl. Biol. 33: 66, 1946.
- Govier & Woods, J. gen. Virol. 13: 127, 1971.
- Hamilton, Ann. appl. Biol. 19: 550, 1932.
- Harrison & Roberts, J. gen. Virol. 10: 71, 1971.
- Kahn & Bartels, Phytopathology 58: 587, 1968.
- Lovisolo & Bartels, Phytopath. Z. 69: 189, 1970.
- Plumb & Vince, J. gen. Virol. 13: 357, 1971.
- Shepherd, Fulton & Wakeman, Phytopathology 59: 219, 1969.
- Watson, Rep. Rothamsted exp. Stn 1967: 123, 1968.
- Watson & Roberts, Proc. R. Soc. B 127: 543, 1939.