Details of DPV and References
DPV NO: 183 September 1977
Family: Unallocated ssRNA- viruses
Genus: Unassigned ssRNA- virus
Species: Orchid fleck virus | Acronym: OFV
Orchid fleck virus
Y. Doi Laboratory of Plant Pathology, Faculty of Agriculture, University of Tokyo, Bunkyo-ku, Tokyo, Japan
M. U. Chang Laboratory of Plant Pathology, Faculty of Agriculture, University of Tokyo, Bunkyo-ku, Tokyo, Japan
K. Yora Laboratory of Plant Pathology, Faculty of Agriculture, University of Tokyo, Bunkyo-ku, Tokyo, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Doi et al. (1969) and
Chang et al. (1973).
Selected synonyms
- Dendrobium virus (Rev. appl. Mycol. 50: 2989)
- Phalaenopsis virus (Rev. appl. Mycol. 51: 426)
-
A virus with non-enveloped, bacilliform particles about 40 x 150 nm in dip preparations and partially purified preparations, but 32-35 x 100-140 nm in thin sections. It is sap-transmissible to several species of orchids and a few dicotyledonous plants, but with difficulty.
Main Diseases
Causes chlorotic or necrotic flecks in species of Cymbidium, Dendrobium, Oncidium and many other kinds of orchids (Fig. 1, Fig. 2).
Geographical Distribution
Japan; probably also in West Germany (Lesemann & Doraiswamy, 1975) and Brazil (Kitajima et al., 1974).
Host Range and Symptomatology
Found naturally in a wide range of orchid species, and transmitted experimentally to species in the Chenopodiaceae, Solanaceae and Aizoaceae. A morphologically similar virus occurs in coffee and citrus (see Relationships). Sap-inoculation is difficult, but successful when the temperature is higher than 30°C.
-
Diagnostic species
- Dendrobium nobile, Cymbidium alexanderi, and other orchid species. Chlorotic
or necrotic spots, sometimes flecks, first on inoculated leaves, and later on upper
leaves. Incubation period is 3-4 weeks. (Fig. 1,
Fig. 2).
- Nicotiana glutinosa, N. tabacum cv. Bright Yellow, White Burley, Xanthi-nc and KY-57 (tobacco). Chlorotic or necrotic local lesions after 2-3 weeks. No systemic symptoms (Fig. 3, Fig. 4).
-
Propagation species
- Cymbidium spp. (e.g. C. alexanderi) are suitable for maintaining cultures, and are good sources of virus for purification.
-
Assay species
- N. tabacum
and N. glutinosa can be used for local lesion assays.
Strains
None reported.
Transmission by Vectors
The vector is not known. Attempts to transmit by using an aphid (Myzus persicae) and a mealybug (Diaspis boisduvallii) were unsuccessful (M. U. Chang, unpublished data).
Transmission through Seed
None reported.
Serology
No information.
Relationships
The bacilliform or bullet-shaped particles of orchid fleck virus superficially resemble those of plant rhabdoviruses, but they are smaller, lack an outer envelope, and are still infective after treatment with detergents (e.g. Triton X-100). Their fine structure is similar to that of the internal component of rhabdoviruses, but they are more labile than particles of rhabdoviruses. In particle morphology and intracellular effects, orchid fleck virus closely resembles Dendrobium virus (Petzold, 1971), Phalaenopsis virus (Lesemann & Begtrup, 1971), coffee ringspot virus (Kitajima & Costa, 1972) and citrus leprosis virus (Kitajima et al., 1972).
Stability in Sap
Crude Cymbidium leaf sap, diluted up to 10-2 with 0.1 M phosphate buffer pH 7.15, and partially purified preparations are infective after 1 day at 6°C.
Purification
Homogenize diseased orchid leaves at 4°C in 0.1 M phosphate buffer pH 7.0 containing 0.1 M sodium diethyldithiocarbamate, 0.1% L-ascorbic acid, 5% Triton X-100 and 0.5% sodium deoxycholate. Centrifuge the homogenate at 5000 rev/min for 15 min. Centrifuge the supernatant fluid at 30,000 rev/min for 2 h, suspending the pellet in 0.1 M phosphate buffer. Repeat two more cycles of differential centrifugation. Sucrose density gradient centrifugation is used for further purification (Chang et al., 1976).
Properties of Particles
None reported.
Particle Structure
Particles are bacilliform or sometimes bullet-shaped, about 40 x 150 nm in partially purified preparations or in dip preparations pre-fixed in 2% osmium tetroxide before negative staining with phosphotungstate (Fig. 6, Fig. 7, Fig. 8, Fig. 10), but 32-35 x 100-140 nm in thin sections (Fig. 5). Particles have no envelope and show a helical structure with a pitch of 4.5 nm (Chang et al., 1976).
Particle Composition
None reported.
Relations with Cells and Tissues
In thin sections of virus-infected tissues, virus particles occur in the nuclei and cytoplasm, and inclusions of low electron density (‘viroplasms’) are also observed in the nuclei. A series of electron micrographs suggests that virus particles are formed in and around the viroplasms and later move to the nuclear envelope. Finally the nuclei deform and the nuclear membrane disrupts to release the virus particles enclosed. In the nuclei, virus particles exist in the chromatin area singly or arranged side-by-side (Fig. 5), showing crystalline arrays in cross sections. Virus particles are commonly found to attach at one end to the inner nuclear membrane. A number of virus particles surrounded by the inner membrane often show an appearance like a spoked wheel (Fig. 9). Some particles appear to associate with or attach to the endoplasmic reticulum. In addition to virus particles, vacuolated or deformed nuclei, swollen or amoeboid chloroplasts, and swollen mitochondria are observed in virus infected cells (Chang et al., 1976).
Figures

Systemic necrotic flecks on infected Cymbidium leaves.

Systemic necrotic flecks on infected Oncidium leaves.
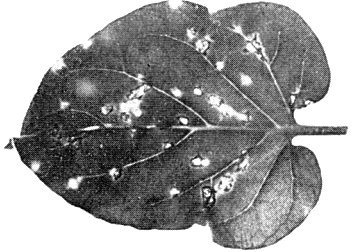
Local chlorotic-necrotic spots on a leaf of N. glutinosa.

Local chlorotic spots on a leaf of N. tabacum cv. Xanthi-nc.

Viroplasm adjacent to nucleolus in the nucleoplasm (N. glutinosa). Ne: nuclear envelope; nu: nucleolus; v: virus; vp: viroplasm. Bar represents 1 µm.

Bullet-shaped particle. Bar represents 50 nm.

Intact bacilliform particle. Bar represents 50 nm.

Virus particles in negatively stained preparation from infected Cymbidium leaves. Bar represents 100 nm.
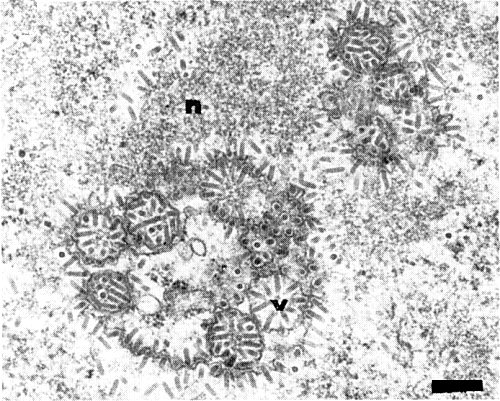
Virus particles surrounded by inner nuclear membrane in perinuclear area, forming ‘spoked wheel’ structures. n: nucleus; v: virus. Bar represents 250 nm.

Virus particles in partially purified preparation. Bar represents 100 nm.
References list for DPV: Orchid fleck virus (183)
- Chang, Arai, Doi & Yora, Ann. Phytopath. Soc. Japan 39: 171, 1973.
- Chang, Arai, Doi & Yora, Ann. Phytopath. Soc. Japan 42: 156, 1976.
- Doi, Toriyama, Yora & Asuyama, Ann. Phytopath. Soc. Japan 35: 388, 1969.
- Kitajima & Costa, Cienciae Cultura 24: 542, 1972.
- Kitajima, Müller, Costa & Yuki, Virology 50: 254, 1972.
- Kitajima, Blumenschei & Costa, Phytopath. Z. 81: 280, 1974.
- Lesemann & Begtrup, Phytopath. Z. 71: 257, 1971.
- Lesemann & Doraiswamy, Phytopath. Z. 83: 27, 1975.
- Petzold, Phytopath. Z. 70: 45, 1971.