Details of DPV and References
DPV NO: 90 June 1972
Family: Alphaflexiviridae
Genus: Potexvirus
Species: Cassava common mosaic virus | Acronym: CsCMV
Cassava common mosaic virus
A. S. Costa Virus Department, Instituto Agronômico, Campinas, S.P., Brazil
E. W. Kitajima Virus Department, Instituto Agronômico, Campinas, S.P., Brazil
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by
Costa (1940).
Selected synonym
- Brazilian common cassava mosaic virus (Rev. appl. Mycol. 44: 312)
An RNA-containing virus with particles about 495 nm long and 15 nm in diameter, belonging to the potato virus X group. No vector is known. Perpetuated in infected cuttings and easily transmitted mechanically, this being considered its natural means of spread. Known from Brazil and Peru.
Main Diseases
Causes a typical mosaic disease with or without leaf malformation in all tested varieties of Manihot esculenta (Fig. 1, Fig. 2). Symptoms are similar to those of African cassava mosaic (Storey & Nichols, 1938), which is caused by a distinct virus. Crop losses induced by virus infection may be 30% or more, but the disease is of minor importance because it is controlled easily by selecting healthy cuttings for propagation, and by roguing.
Geographical Distribution
Occurs in various parts of Brazil where cassava is grown. Also found in quarantined cassava plants at Bogota, Colombia, in cuttings introduced from Peru (E. W. Kitajima, unpublished).
Host Range and Symptomatology
Infects species from at least six dicotyledonous families (Amaranthaceae, Chenopodiaceae, Compositae, Euphorbiaceae, Malvaceae and Solanaceae). Easily transmitted mechanically.
Diagnostic species
- Chenopodium album, C. amaranticolor
(Fig. 3), C. murale and C. quinoa develop small chlorotic or necrotic local lesions, but no systemic infection.- Gossypium hirsutum (cotton) develops large, irregular local lesions and systemic
vein-clearing, then recovers.
- Gomphrena globosa shows local necrotic lesions usually larger than those caused by potato virus X, but fewer; no systemic infection.
- Ricinus communis (castor bean) shows systemic mottling in the early stages of infection, but symptoms become masked as infected plants mature.
- Gomphrena globosa shows local necrotic lesions usually larger than those caused by potato virus X, but fewer; no systemic infection.
Propagation species
- Manihot esculenta
(cassava) cuttings are good plants in which to maintain the virus. Euphorbia prunifolia is a symptomless carrier, but contains a high virus concentration and is a good source plant for purification.Assay species
- Chenopodium
species are good local lesion assay hosts.
Strains
Strains differing in virulence on cassava have been isolated, but no detailed studies of them have been made.
Transmission by Vectors
Tests with aphids (Aphis gossypii, Myzus persicae), white flies (Bemisia tabaci), thrips (Scirtothrips manihoti) and mites (Mononychus bondari) gave negative results.
Transmission through Seed
No virus was detected in over 100 cassava seedlings grown from seed of mosaic-diseased plants.
Serology
Low-titred antiserum (1/64 in precipitin tests) was obtained by injecting rabbits intravenously with clarified sap of infected Euphorbia prunifolia (Silva, 1962). Using partially purified virus from infected cassava, plus Freund’s incomplete adjuvant, an antiserum was obtained with a titre of 1/4096 in tube precipitin tests (Kitajima et al., 1965).
Relationships
In physical and morphological properties the cassava common mosaic virus resembles viruses of the potato virus X group, but its modal length was consistently 17 nm shorter than that of potato virus X (Kitajima et al., 1965). There was no serological cross-reaction between the two viruses (Kitajima et al., 1963, 1965).
Stability in Sap
Most virus infectivity is destroyed when sap of infected cassava or Euphorbia prunifolia plants is heated at 65°C for 10 min; some infectivity is still detectable after 10 min at 70°C, but not at higher temperatures. In sap of E. prunifolia infectivity is lost after dilution between 10-5 and 10-6, but not after 128 days’ storage at room temperature (A. S. Costa, unpublished).
Purification
Sap of infected Euphorbia prunifolia was clarified by adding n-butanol to 8% (v/v), followed by low speed centrifugation, filtration through absorbent cotton and 2-4 cycles of high and low speed centrifugation (Silva, Kitajima & Oliveira, 1963). Partial purification was obtained by clarifying cassava sap with an ether-carbon tetrachloride mixture, followed by one cycle of high and low speed centrifugation (Kitajima et al., 1965).
Properties of Particles
Not determined.
Particle Structure
Particles are flexuous filaments c. 495 nm long, 15 nm in diameter (Fig. 4). No internal channel was seen in intact or broken particles of the virus mounted in 1% phosphotungstate (Kitajima et al., 1965).
Particle Composition
Optical density measurements of purified preparations indicated the virus to be a nucleoprotein and tests with orcinol and diphenylamine suggest that its nucleic acid is RNA. Absorbance data at 260/280 nm suggest that about 5% of the particle weight is RNA (Silva et al., 1963).
Relations with Cells and Tissues
Virus inclusions (Fig. 5) occur in the cytoplasm of all leaf tissues of infected cassava plants, except tracheids and sieve tubes. They consist of fibrous masses containing filamentous particles of indeterminate length and 10-15 nm in diameter, running more or less parallel, but not in a definite arrangement. Virus inclusions are variable in size, generally elongated, up to 15-20 µm long and 3-5 µm wide as seen in sections. They are more frequent in yellow leaf areas. No major changes in cell components were associated with virus infection, except for abnormal accumulation of starch in the chloroplasts (Kitajima & Costa, 1966).
Notes
Cassava common mosaic virus can be easily distinguished from potato virus X on the basis of host range and symptoms produced on several solanaceous hosts. It may be confused with malva veinal necrosis virus (Costa & Kitajima, 1970) but the latter virus differs in having somewhat longer particles and a more restricted host range. The cassava common mosaic virus from Brazil differs from African cassava mosaic virus (Storey & Nichols, 1938) because it is easily transmitted by inoculation of sap and is not transmitted by white flies; also no elongated particles were found in cassava plants infected with the African virus (Kitajima & Costa, 1964). The causal agent of cassava witches’ broom (Silberschmidt & Campos, 1944) has been listed as a strain or suspected strain of African cassava mosaic virus (Rev. appl. Mycol. 35, Suppl.: 12) and later also considered a synonym of cassava common mosaic virus of Brazil (Martyn, 1968). There has never been good evidence for this because cassava witches’ broom is a disease of the yellows type; it is now known to be associated with mycoplasma infection (Kitajima & Costa, 1970).
Figures

Infected cassava leaf showing mosaic and malformation.

Infected cassava leaf showing mosaic symptoms only.

Inoculated leaf of Chenopodium amaranticolor showing local lesions.

Virus particles from cassava sap in 1% phosphotungstate. Bar represents 200 nm.
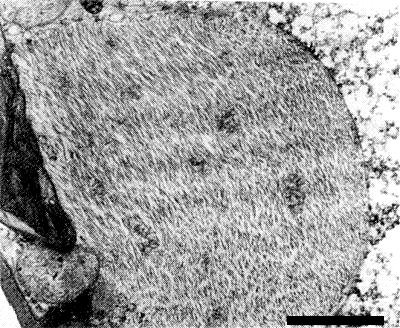
Virus inclusion in a leaf parenchyma cell. Bar represents 1 µm.
References list for DPV: Cassava common mosaic virus (90)
- Costa, Jorn. Agron. 3: 239, 1940.
- Costa & Kitajima, Bragantia 29: li, 1970.
- Kitajima & Costa, E. Afr. agric. For. J. 30: 28, 1964.
- Kitajima & Costa, Bragantia 25: xxiii, 1966.
- Kitajima & Costa, Ciênc. Cult. 22: 351, 1970.
- Kitajima, Silva, Costa & Carvalho, Ciênc. Cult. 15: 305, 1963.
- Kitajima, Wetter, Oliveira, Silva & Costa, Bragantia 24: 247, 1965.
- Martyn, Phytopath. Pap. 9, 204 pp., 1968.
- Silberschmidt & Campos, Arq. Inst. biol. S. Paulo 15: 1, 1944.
- Silva, Bragantia 21: xcix, 1962.
- Silva, Kitajima & Oliveira, Ciênc. Cult. 15: 304, 1963.
- Storey & Nichols, Ann. appl. Biol. 25: 790, 1938.