Details of DPV and References
DPV NO: 34 October 1970
Family: Reoviridae
Genus: Phytoreovirus
Species: Wound tumor virus | Acronym: WTV
Wound tumor virus
L. M. Black Dept. of Botany, University of Illinois, Urbana, Illinois 61801, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Black (1944).
Selected synonyms
- Clover wound tumor virus (Rev. appl. Mycol. 47: 461)
- Clover big vein virus (Rev. appl. Mycol. 23: 490)
- Aureogenus magnivena (Rev. appl. Mycol. 23: 490)
- Trifoliumvirus nervicrassans (Rev. appl. Mycol. 38: 677)
- Clover big vein virus (Rev. appl. Mycol. 23: 490)
-
An isometric virus with particles c. 70 nm in diameter containing double-stranded RNA. Only six instances of sap transmission to plants have been reported but the virus multiplies in certain agallian leafhoppers and is transmitted by them to a wide range of host plants.
Main Diseases
Wound tumor virus (WTV) was found in the vector. It has not been found causing disease in any plant in the field.
Geographical Distribution
Collected once in leafhoppers in the vicinity of Washington, DC, USA.
Host Range and Symptomatology
Species in at least 20 families of dicotyledonous plants are susceptible (Black, 1945). Vein enlargement, not vein-clearing, is the first systemic symptom. Root tumours are characteristic (Fig. 6); stem tumours occur in a few species (Fig. 5). The genetic constitution of individual plants (Black, 1951), wounds (Black, 1965) and plant hormones (Black & Lee, 1957) influence the initiation of tumours in infected plants; in sweet clover, tumours are first detectable in the pericycle at the 4th or 5th incipient lateral root (Kelly & Black, 1949; Lee, 1955).
-
Diagnostic species
- Trifolium incarnatum
(crimson clover). Characteristic irregular vein enlargement (Fig. 1, Fig. 2, Fig. 3). - Melilotus officinalis and M. alba (sweet clovers), and Rumex
acetosa (cultivated sorrel). Variable root tumour response among individual
plants. M. officinalis clone C10 develops many root tumours
(Black, 1951).
- Medicago sativa cv. Grimm (alfalfa) is immune.
-
Propagation species
- Melilotus officinalis
clone C10, by propagation of infected cuttings. Virus is most concentrated in the tumours, about 1012 virions/g.Assay species
- Trifolium incarnatum
is preferred for testing viruliferous insects. Virus can be assayed by injection into vectors (Maramorosch, Brakke & Black, 1949) which are then tested for transmission of virus to plants or for production of virus antigens (Whitcomb, 1964; Reddy & Black, 1966). Cell monolayer cultures from Agallia constricta provide the best assay; a preparation containing c. 106 virions/ml or more is needed to cause infection (Gamez & Chiu, 1968). Infected cells are detected by staining with fluorescent antibody; the relation between virus concentration in inoculum and number of infected cells is linear (Chiu, Reddy & Black, 1966; Chiu & Black, 1967, 1969).
Strains
Growth of the virus in sweet clover cuttings over a period of years without transmission through vectors results in strains (‘subvectorial’) that are poorly transmissible and, eventually, in strains (‘exvectorial’) that are not transmissible by the vector (Black, 1969). The subvectorial strains have low specific infectivities for vector cell monolayers and the exvectorial strains have no infectivity. Subvectorial strains produce less virus and more soluble antigen in sweet clover tumours than does the wild (or ‘vectorial’) strain. The strain, exvectorial 49, can no longer be purified from tumours (Whitcomb & Black, 1969; Reddy & Black, 1969; Gamez & Black, 1969).
Transmission by Vectors
The virus is transmitted by certain agallian leafhoppers, especially Agallia constricta and Agalliopsis novella (Fig. 4) (Black, 1944), in which it multiplies (Black & Brakke, 1952; Brakke, Vatter & Black, 1954). Aceratagallia sanguinolenta does not transmit. At 27°C the virus has a minimum incubation period in the vectors of about 2 weeks (Maramorosch, 1950) during which it infects the intestinal tract, haemolymph and salivary glands in sequence (Sinha, 1965, 1967). It reaches a peak concentration of c. 109 virions per leafhopper (i.e. per mg) in about 4 weeks (Reddy & Black, 1966; Gamez & Black, 1968). Frequency of transovarial transmission in A. constricta is genetically controlled; this control is independent of the genetic variation in ability to transmit potato yellow dwarf virus (Black, 1953; Nagaraj & Black, 1962; Sinha & Shelley, 1965).
Transmission through Seed
No information.
Transmission by Dodder
No information.
Serology
Antisera react in precipitin, complement-fixation or neutralization tests with virus or soluble antigen both in plant and in insect extracts (Black & Brakke, 1954). The precipitin ring test or ring-time test has been used to measure the increase of soluble antigen (Whitcomb & Black, 1961a, 1961b) and, indirectly, virus infectivity (Reddy & Black, 1966). Fluorescent antibody may be used in haemolymph smear tests on individual insects (Nagaraj, Sinha & Black, 1961) without killing them (Sinha, Reddy & Black, 1964). The minimum concentrations of virions detectable by the ring test and by the passive haemagglutination test were 7 x 109/ml and 2.5 x 108/ml, respectively (Gamez & Black, 1967).
Relationships
Antisera to vectorial virus strains react with virus and soluble antigen of subvectorial and exvectorial strains. WTV closely resembles the reoviruses of vertebrates in particle morphology and in having several double-stranded RNA components and a RNA polymerase. However, sensitive passive haemagglutination tests failed to show relationships between WTV and three strains of reovirus (Gamez, Black & MacLeod, 1967). Virion morphology, nucleic acid structure and intracellular behaviour suggest that rice dwarf, maize rough dwarf and sugarcane Fiji disease viruses may have affinities with WTV.
Stability in Sap
In vector extracts the thermal inactivation point (10 min) is 50-60°C; infectivity survived in dried extracts for at least 1 year at 0°C (Brakke, Maramorosch & Black, 1953). At 37°C WTV is most stable between pH 6.60 and 6.61 (Kimura & Black, unpublished).
Purification
From root tumors of sweet clover clone C10: tumor production is favoured by a daily cycle of 16 h of light at 20°C and 8 h of darkness at 17°C (Black, 1965). Purified by density gradient centrifugation and zonal electrophoresis in density gradients (Brakke et al., 1954). Treatment with carbon tetrachloride and Freon aids purification (Kimura & Black, unpublished). Virus can also be purified from vectors.
Properties of Particles
The particles seem all of one type; the dilution curve indicates one particle causes infection (Chiu & Black, 1969).
Sedimentation coefficient (s20,w) at infinite dilution: c. 510 S (Black, Hills & Markham, 1962) or 514 ± 10 S (Kalmakoff, Lewandowski & Black, 1969).
Molecular weight (daltons): 68 ± 2 x 106 (Kalmakoff et al., 1969).
Absorbance at 260 nm (1 cm light path): 1.0 for the hydrolysed RNA from suspensions containing 10 x 1011 virions/ml (Ahmed & Black, unpublished); for the virus, an absorbance of 1.0 (after correction for light scattering) = 8.8 x 1011 virions/ml (Kimura & Black, unpublished).
A260/A280: c. 1.55; A260/A243: c. 1.16 (Black et al., 1962).
Particle Structure
Particles are icosahedral, c. 70 nm in diameter (Fig. 7). The virus seems to have a membrane supporting 32 capsomeres and enclosing the double-stranded RNA (Black et al., 1962; Streissle & Granados, 1968; Kimura & Black, unpublished).
Particle Composition
RNA: 22% of the weight of the particle, double-stranded (Fig. 8), containing 38% G + C (Bils & Hall, 1962; Black & Markham, 1963; Gomatos & Tamm, 1963; Kleinschmidt et al., 1964; Tomita & Rich, 1964). Each particle contains about 15-16 x 106 daltons of nucleic acid (Black & Markham, 1963; Kalmakoff et al., 1969; Wood & Streissle, 1970), which when separated from the particle is found to consist of 12 molecules (Kalmakoff et al., 1969) or 15 molecules (Wood & Streissle, 1970), mostly of different sizes. Isolated RNA is not infective.
Protein: 78% of the particle weight. The number and amounts of different proteins in the particles is not known. A RNA polymerase is associated with the virus (D. R. Black & C. A. Knight, personal communication).
Relations with Cells and Tissues
In plants, virus antigen is mostly restricted to the abnormal phloem where it is concentrated in spherical bodies in the cytoplasm (Nagaraj & Black, 1961). Perhaps these are the inclusions of Littau & Black (1952) or the viroplasms of Shikata & Maramorosch (1965). Viruliferous vectors show no macroscopic evidence of disease. Virions occur in the cytoplasm but not in the nucleus of many kinds of vector cells, some of which show cytopathic changes (Hirumi, Granados & Maramorosch, 1967; Maramorosch et al., 1968; Shikata & Maramorosch, 1967a, 1967b). Fluorescent antibodies stain the cytoplasm but not the nucleus of infected vector cells (Chiu et al., 1970).
Notes
Tobacco club root (Rev. appl. Mycol. 27: 101) was listed as another disease induced by wound tumor virus (Rev. appl. Mycol. 47: 2420). The evidence is to the contrary (Selsky, 1961).
Figures

Crimson clover, Trifolium incarnatum. Upper surface of infected leaf showing depressed veins.

Crimson clover, Trifolium incarnatum. Lower surface of infected leaf showing irregular vein enlargement.

Crimson clover, Trifolium incarnatum. Smooth upper surface of healthy leaf (except for the mid-vein, veins on the lower surface are no more prominent).
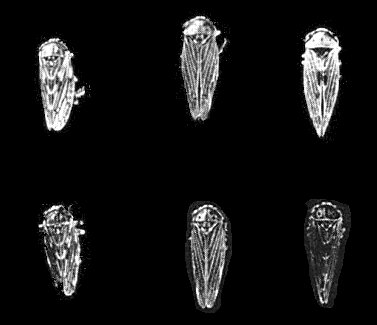
Agallian leafhoppers, (upper row) females, (lower row) males. (Left to right) Aceratagallia sanguinolenta (not a vector), Agallia constricta (a vector) and Agalliopsis novella (a vector).
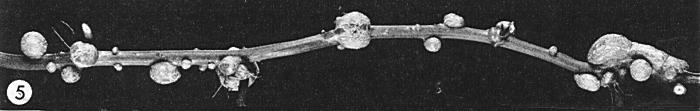
Stem tumours on infected sweet clover (Melilotus sp.).

Young infected root of Rumex acetosa. Tumours tend to arise above, below, or at the side, but not opposite, the emergence points of lateral roots.

Virions in phosphotungstate. Bar represents 150 nm. (Courtesy of I. Kimura.)

Double-stranded RNA filaments. Bar represents 150 nm. (Courtesy of Kleinschmidt et al., 1964.)
References list for DPV: Wound tumor virus (34)
- Bils & Hall, Virology 17: 123, 1962.
- Black, Proc. Am. phil. Soc. 88: 132, 1944.
- Black, Am. J. Bot. 32: 408, 1945.
- Black, Am. J. Bot. 38: 256, 1951.
- Black, Phytopathology 43: 9, 1953.
- Black, In Encyclopedia of Plant Physiology, Springer-Verlag,New York, pp. 236-266, 1965.
- Black, A. Rev. Phytopath. 7: 73, 1969.
- Black & Brakke, Phytopathology 42: 269, 1952.
- Black & Brakke, Phytopathology 44: 482, 1954.
- Black, Hills & Markham, J. gen. Microbiol. 28: 1, 1962.
- Black & Lee, Virology 3:146, 1957.
- Black & Markham, Neth. J. Pl. Path. 69: 215, 1963.
- Brakke, Maramorosch & Black, Phytopathology 43: 387, 1953.
- Brakke, Vatter & Black, Brookhaven Symp. Biol. 6: 137, 1954.
- Chiu & Black, Nature, Lond. 215: 1076, 1967.
- Chiu & Black, Virology 37: 667, 1969.
- Chiu, Liu, MacLeod & Black, Virology 40: 387, 1970.
- Chiu, Reddy & Black, Virology 30: 562, 1966.
- Gamez & Black, Nature, Lond. 215: 173, 1967.
- Gamez & Black, Virology 34: 444, 1968.
- Gamez & Black, A. Rev. Phytopath. 7: 92, 1969.
- Gamez, Black & MacLeod, Virology 32: 163, 1967.
- Gamez & Chiu, Virology 34: 356, 1968.
- Gomatos & Tamm, Proc. natn. Acad. Sci. U.S.A. 50: 878, 1963.
- Hirumi, Granados & Maramorosch, J. Virol. 1: 430, 1967.
- Kalmakoff, Lewandowski & Black, J. Virol. 4: 851,1969.
- Kelly & Black, Am. J. Bot. 36: 65, 1949.
- Kleinschmidt, Dunnebacke, Spendlove, Schaffer & Whitcomb, J. molec. Biol. 10: 282, 1964.
- Lee, Virology 1: 152, 1955.
- Littau & Black, Am. J. Bot. 39: 87, 1952.
- Maramorosch, Phytopathology 40: 1071, 1950.
- Maramorosch, Brakke & Black, Science, N.Y. 110: 162, 1949.
- Maramorosch, Shikata, Hirumi & Granados, Natn. Cancer Inst. Monogr. 31: 493, 1968.
- Nagaraj & Black, Virology 15: 289, 1961.
- Nagaraj & Black, Virology 16: 152, 1962.
- Nagaraj, Sinha & Black, Virology 15: 205, 1961.
- Reddy & Black, Virology 30: 551, 1966.
- Reddy & Black, A. Rev. Phytopath. 7: 88, 1969.
- Selsky, Phytopathology 51: 581, 1961.
- Shikata & Maramorosch, Virology 27: 461, 1965.
- Shikata & Maramorosch, Virology 32: 363, 1967a.
- Shikata & Maramorosch, J. Virol. 1:1052, 1967b.
- Sinha, Virology 26: 673, 1965.
- Sinha, Virology 31: 746, 1967.
- Sinha, Reddy & Black, Virology 24: 666, 1964.
- Sinha & Shelley, Phytopathology 55: 324, 1965.
- Streissle & Granados, Archiv. Virusforsch. 25: 369, 1968.
- Tomita & Rich, Nature, Lond. 201: 1160, 1964.
- Whitcomb, Virology 24: 488, 1964.
- Whitcomb & Black, Virology 15: 136, 1961a.
- Whitcomb & Black, Virology 15: 507, 1961b.
- Whitcomb & Black, A. Rev. Phytopath. 7: 86, 1969.
- Wood & Streissle, Virology 40: 329, 1970.