Details of DPV and References
DPV NO: 72 October 1971
Family: Reoviridae
Genus: Fijivirus
Species: Maize rough dwarf virus | Acronym: MRDV
Maize rough dwarf virus
O. Lovisolo Laboratorio di Fitovirologia applicata del CNR, 10135 Torino, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Biraghi (1949,
1952),
Harpaz (1959).
Reviewed by
Lovisolo (1971).
Selected synonyms
- Nanismo ruvido del mais (Rev. appl. Mycol. 32: 249)
- Rauhverzwergung des Maises (Rev. appl. Mycol. 44: 3034)
- (Maize rough dwarf and stripe disease (Rev. appl. Mycol. 44: 3034) is probably caused by maize rough dwarf virus in combination with the mycoplasm associated with aster yellows disease)
- Rauhverzwergung des Maises (Rev. appl. Mycol. 44: 3034)
-
A virus with isometric particles c. 70 nm in diameter. It has not been transmitted by rubbing methods but is transmitted to graminaceous plants by planthoppers, in which it multiplies. It occurs in Europe and the Middle East where it damages maize.
Main Diseases
Maize (Zea mays) infected when young is severely dwarfed (Fig. 1) and there is practically no grain yield. Typically, small enations occur on the lower surfaces of the leaves (Fig. 2) and sometimes also on the sheaths of leaves and ears (the enations are rough to the touch, hence the name of the disease). Adventitious roots have very few secondary roots, and are short and thick with longitudinal cracks. Ears, if produced, are very small and their distal portions are atrophied. For further details of the reactions of different varieties see Grancini (1958, 1962). Plants infected when older show little or no dwarfing, and the only noticeable symptoms are the typical enations.
Geographical Distribution
Occurs in Europe (Italy, Switzerland, Spain, France, Yugoslavia, Czechoslovakia) and in Israel. The disease has only been recorded where the vector Laodelphax striatellus is present (see Commonwealth Institute of Entomology, Distribution Maps of Pests, Series A, Map No. 201, June 1965).
Host Range and Symptomatology
The only known natural hosts are maize, Digitaria sanguinalis (Luisoni & Conti, 1970) and Echinochloa crus-galli (M. Conti, unpublished). There may be more natural hosts in the Gramineae, because 16 species (mainly in the tribes Paniceae and Hordeae) have been infected experimentally by vectors. The virus has not been transmitted by rubbing methods; transmission by needle-prick is possible (Harpaz, 1959) but difficult. In experimentally infected wheat, oats and barley (Fig. 3) the symptoms are dwarfing, early and excessive tillering, and stem stunting (Vidano, Lovisolo & Conti, 1966a, 1966b; Lovisolo, Vidano & Conti, 1966).
-
Diagnostic species
- Zea mays
(maize), Laguna Elite, Nevé Yaar 22 and American yellow dent hybrids such as Wisconsin 641 AA. Young seedlings are infected at the coleoptile stage using Laodelphax striatellus as vector (Harpaz et al., 1965). Leaves of infected plants first become darker green, then develop numerous minute enations on the veins. Growth is decreased and severe dwarfing results.Propagation species
- Maize, barley, wheat and oats are suitable plants for maintaining cultures. Maize roots of American yellow dent hybrids such as Wisconsin 641 AA are the most convenient source of virus for purification.
-
Assay species
- Maize cv. Wisconsin 641 AA or similar hybrids are useful systemic hosts in infectivity tests using planthoppers as vectors. No local lesion hosts are known. The infectivity of clarified plant sap and vector haemolymph (Harpaz & Klein, 1969), and of crude extracts from maize enations (M. Conti, unpublished), may be tested by injecting them into vector insects and allowing the insects to feed on maize test plants, but purified virus preparations tested in this way have low infectivity (M. Conti, unpublished).
Strains
According to Harpaz & Klein (1969) there are two distinct virus strains (‘dwarfing strain’ and ‘non-dwarfing strain’), characterized by different symptoms and different relations with vectors and host plants. These findings, however, require further study and I. Harpaz (pers. comm.) does not exclude other explanations.
Transmission by Vectors
Transmitted by planthoppers of the family Delphacidae. The only natural vector so far known is Laodelphax striatellus. The minimum durations of the acquisition, incubation and inoculation periods are respectively: 1 day, 10-15 days and 5 h (Harpaz et al., 1965; Conti, 1966). The virus multiplies in the vector (Vidano, 1970) and seems to be pathogenic towards it. Only a small proportion of the offspring of infected females was infected (Harpaz & Klein, 1969) because most of the infected progeny die during embryonic development (I. Harpaz, pers. comm.). Experimentally, Javesella pellucida (Harpaz et al., 1965) and Sogatella vibix can acquire and transmit the virus between plants, and Delphacodes propinqua can transmit the virus to plants after infective sap is injected into its thoracic cavity (Harpaz & Klein, 1969).
Many factors affect the amount of infection of maize crops (Harpaz, 1961; Klein & Harpaz, 1969; Grancini & Corte, 1969), one of the most important being that L. striatellus does not breed on maize, and only feeds on this plant occasionally.
Transmission through Seed
None found.
Transmission by Dodder
Not transmitted by Cuscuta eigii (Harpaz, 1959).
Serology
The particles of 55-60 nm diameter (see Particle structure) are moderately immunogenic. Antisera have been obtained with titres up to 1/2048 (Wetter et al., 1969) when tested either in agar gel-diffusion or in slide precipitin tests. Virus from infected plants and planthoppers is antigenically indistinguishable.
Relationships
Maize rough dwarf virus shares many properties with animal viruses of the reovirus group. Among the plant viruses of this type, it shares most features with rice black-streaked dwarf virus, pangola stunt virus (Kitajima & Costa, 1970) and sugar-cane Fiji disease virus (Teakle & Steindl, 1969). A close serological relationship between maize rough dwarf virus and rice black-streaked dwarf virus has been found by agar gel-diffusion, ring-precipitin and slide-precipitin tests (joint work in progress between the Laboratorio di Fitovirologia applicata, Turin, Italy, and the Botanical Institute of the Faculty of Agriculture, Sapporo, Japan). For this work preparations of the 55-60 nm diameter particles were used (see Particle structure). No serological affinity has been detected between maize rough dwarf virus and wound tumor virus (Wetter et al., 1969).
Stability in Sap
There is no information on the stability of the virus in sap, but partially purified preparations of the virus were still serologically active after 37 days at room temperature, after 10 min at 70° but not 80°C and after 3 days at -25°C; serological activity diminished above pH 9 (Wetter et al., 1969).
Purification
Washed roots are extracted in the presence of 0.03 M Na2HPO4 + 0.01 M Na2SO3 + 0.001 M Na-ethylenediamine-tetraacetate, filtered through diatomaceous earth, and the virus purified by two cycles of differential centrifugation with one or two intervening ultracentrifugations in sucrose density gradients (Wetter et al., 1969). Alternatively, the extract may be clarified by shaking it with an equal amount of chloroform and then breaking the emulsion by low speed centrifugation; this is the most suitable method when purifying from stems and leaves, and is also the only method suitable for purification from planthoppers (Wetter et al., 1969). Better preparations were obtained from maize roots than from leaves and the final preparations always contain only isometric particles of 55-60 nm diameter (Fig. 6). These are probably particles without an outer shell (see Particle Structure).
Properties of Particles
For the 55-60 nm particles: sedimentation coefficient (s20,w): c. 400 S; A260/A280: c. 1.5 (P. Redolfi, H. L. Paul & S. Pennazio, pers. comm.).
Particle Structure
The structure of the particles is probably similar to that of the animal-infecting reoviruses, which have a core of about 40-50 nm, surrounded by an inner and an outer shell. In sections of maize rough dwarf virus-infected plant and planthopper tissues, and in crude extracts (Fig. 4, Fig. 5) from neoplastic tissues treated with uranyl acetate (Lovisolo, 1967) or uranyl formate (D. Lesemann, pers. comm.), many particles are isometric, c. 70 nm in diameter. In purified preparations or in crude extracts of neoplastic tissues treated with neutral 2% potassium phosphotungstate (Lovisolo, 1967), particles only 55-60 nm in diameter are found and these are probably the virus particles devoid of the outer shell (Fig. 6).
Particle Composition
The nucleic acid of the virus is RNA (P. Redolfi, A. Cantisani & S. Pennazio, pers. comm.).
Relations with Cells and Tissues
Ultrathin sections of infected maize and wheat show numerous virus particles in the leaf and root cells of neoplastic phloem and also in mesophyll cells near the phloem (Gerola & Bassi, 1966; Gerola et al., 1966b; Lovisolo & Conti, 1966; Kislev, Harpaz & Klein, 1968; A. Appiano & O. Lovisolo, unpublished). In plants the virus occurs in ‘viroplasms’ (40-50 nm particles, enclosed in dense masses of fine fibrils), as aggregates of particles in crystalline arrays (Fig. 7) and as 70 nm particles scattered in the cytoplasm. The virus particles sometimes become one of the main components of the cell. Particles of 70 nm diameter may occur in single rows within straight tubules of various lengths (Fig. 5). In ultrathin sections of the vector Laodelphax striatellus (Gerola et al., 1966a; Vidano, 1966, 1967, 1968, 1970) virus particles are found in fat bodies, salivary glands, mycetome, gut, muscles, epidermis, tracheae, ovarioles and brain. Vidano (1970) recorded eight types of virus-induced structures in the vector (some of which have not been reported in plants: e.g. granular microtubules and phagocytic vesicles) which he considers to be stages in the growth of the virus.
Notes
Mild enation symptoms caused by maize rough dwarf virus in maize might be confused with ephemeral vein-swellings caused by leafhopper saliva (Harpaz, 1966) but the virus can be readily diagnosed by looking for particles in crude extracts from the swellings by electron microscopy (Lovisolo, 1967). The virus causes symptoms in oats, wheat and barley very similar to those induced by oat sterile dwarf virus, but it has never been found infecting these plants in the field. Epidemiological studies have led to satisfactory control measures: e.g. delayed sowing (Harpaz, 1961) and killing the vector on the weedy grass borders surrounding the fields about 20-30 days before sowing maize (Grancini & Corte, 1969). Old-world maize cultivars are less susceptible than new American dent hybrids (Harpaz, 1961; Grancini, 1962).
Acknowledgements
Photographs: Laboratorio di Fitovirologia applicata, Torino; Fig. 5 courtesy D. Lesemann.
Figures

Maize plants, (left) healthy, (right) infected, 45 days after inoculation.

Part of infected maize plant with typical enations, 49 days after inoculation.
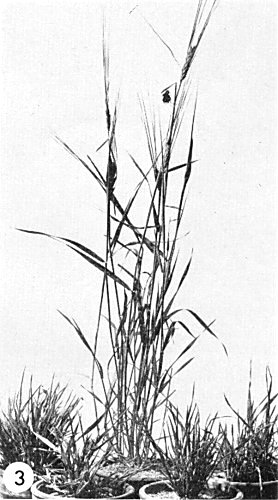
Barley, showing four infected plants, 6 months after inoculation, surrounding a normal plant.
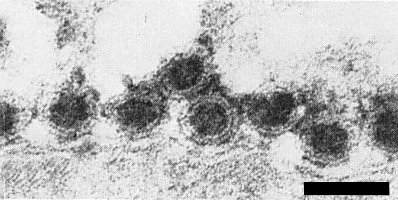
Virus particles from enations; treated with uranyl acetate. Bar represents 100 nm.
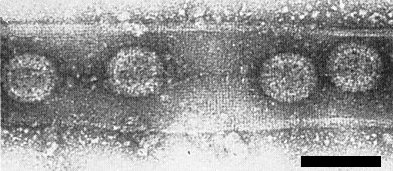
Virus particles within a straight tubule from enations; treated with uranyl formate. Bar represents 100 nm.

Particles (55-60 nm) from a purified preparation in phosphotungstate. Bar represents 100 nm.

Electron micrograph of a thin section through an infected maize root cell showing virus particles in crystalline aggregates (Va) and ‘viroplasm’ (Vp). Bar represents 250 nm.
References list for DPV: Maize rough dwarf virus (72)
- Biraghi, Notiz. Mal. Piante 7: 1, 1949.
- Biraghi, Annali Sper. agr. 6: 1043, 1952.
- Conti, Annali Fac. Sci. agr. Univ. Torino 3: 337, 1966.
- Gerola & Bassi, Caryologia 19: 13, 1966.
- Gerola, Bassi, Lovisolo & Vidano, Phytopath. Z. 56: 97, 1966a.
- Gerola, Bassi, Lovisolo & Vidano, Caryologia 19: 493, 1966b.
- Grancini, Maydica 3: 67, 1958.
- Grancini, Maydica 7: 17, 1962.
- Grancini & Corte, Maydica 14: 79, 1969.
- Harpaz, Nature, Lond. 184: 77, 1959.
- Harpaz, Pl. Prot. Bull. F.A.O. 9: 144, 1961.
- Harpaz, Maydica 11: 18, 1966.
- Harpaz & Klein, Entomologia exp. appl. 12: 99, 1969.
- Harpaz, Vidano, Lovisolo & Conti, Atti Accad. Sci., Torino 99:885, 1965.
- Kislev, Harpaz & Klein, Acta phytopath. Acad. Sci. hung. 3: 1, 1968.
- Kitajima & Costa, Proc. VII Congrès Internat. Microscopie Electr., Grenoble 3: 323, 1970.
- Klein & Harpaz, Z. angew. Ent. 64: 39, 1969.
- Lovisolo, Atti Accad. Sci., Torino 101: 615, 1967.
- Lovisolo, TagBer. dt. Akad. LandwWiss. Berl. 115: (in press), 1971.
- Lovisolo & Conti, Atti Accad. Sci., Torino 100: 63, 1966.
- Lovisolo, Vidano & Conti, Atti Accad. Sci., Torino 100:351, 1966.
- Luisoni & Conti, Phytopath. Mediterranea 9: 102, 1970.
- Teakle & Steindl, Virology 37: 139, 1969.
- Vidano, Atti Accad. Sci., Torino 100: 731, 1966.
- Vidano, Atti Accad. Sci., Torino 101: 717, 1967.
- Vidano, Atti Accad. Sci., Torino 102: 641, 1968.
- Vidano, Virology 41: 218, 1970.
- Vidano, Lovisolo & Conti, Atti Accad. Sci., Torino 100: 125, 1966a.
- Vidano, Lovisolo & Conti, Atti Accad. Sci., Torino 106: 699, 1966b.
- Wetter, Luisoni, Conti & Lovisolo, Phytopath. Z. 66: 197, 1969.