Details of DPV and References
DPV NO: 258 July 1982
Family: Potyviridae
Genus: Potyvirus
Species: Tobacco etch virus | Acronym: TEV
This is a revised version of DPV 55
Tobacco etch virus
D. E. Purcifull Department of Plant Pathology, Institute of Food & Agricultural Sciences, University of Florida, Gainesville, Florida 32611, USA
E. Hiebert Department of Plant Pathology, Institute of Food & Agricultural Sciences, University of Florida, Gainesville, Florida 32611, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Blakeslee (1921) and Johnson (1930).
- Selected synonyms
- Datura virus Z (Rev. appl. Mycol. 1: 74)
- Marmor erodens (Rev. appl. Mycol. 28: 514)
- Nicotiana virus 7 (Rev. appl. Mycol. 17: 52)
- Tobacco virus 13 (Rev. appl. Mycol. 17: 52)
- Tomato etch virus (Rev. appl. Mycol. 20: 85)
- Marmor erodens (Rev. appl. Mycol. 28: 514)
- A virus with flexuous filamentous particles 730 nm long and 12-13 nm in diameter containing single-stranded RNA. It induces the formation of crystalline nuclear inclusions and cytoplasmic, cylindrical (pinwheel) inclusions in host cells. It is transmitted mechanically, and by several aphid species in a non-persistent manner. It infects a wide range of experimental hosts and causes diseases of several solanaceous crops in North and South America.
Main Diseases
The severity of symptoms in cultivated plants depends greatly on the host species and cultivar and on virus strain. In tobacco (Nicotiana tabacum), the leaves are narrowed and show mottling and necrotic etching (Johnson, 1930); symptoms and effects on yield can be more severe with Burley tobaccos than with flue-cured types (Stover, 1951; Gooding, 1970; Gooding & Ross, 1970). Symptoms in pepper (Capsicum annuum) include mottling, mosaic and distortion of leaves, distortion of fruit, and stunting (Johnson, 1930; Zitter, 1971); however, resistant cultivars have been developed (Cook et al., 1976; Cook, Zitter & Ozaki, 1977). Infected tomato plants are stunted, and their leaves are mottled and distorted (Johnson, 1930); yield reductions of over 25% were noted (Debrot, 1976).
Jimson weed (Datura stramonium) was the first reported natural host of the virus (Blakeslee, 1921; Chester, 1937); the leaves show mottling, distortion and vein banding, and the capsular spines may be reduced or absent. Cassia obtusifolia (= C. tora), a leguminous weed, is a natural host of the virus in North and South America (Anderson, 1954; Debrot, 1976); systemic symptoms include necrosis, mottling, distortion, stunting and reduced seed production, but the virus may be distributed erratically within the plant (Demski, 1979). Other natural hosts include Cirsium vulgare, Chenopodium album, Linaria canadensis, Physalis spp. and Solanum spp. (Anderson, 1959; Weinbaum & Milbrath, 1976; Debrot, 1976).
Geographical Distribution
Common in North and South America. Reported from Canada, USA (inc. Hawaii), Mexico, Puerto Rico and Venezuela.
Host Range and Symptomatology
Readily transmissible by mechanical inoculation. Over 120 species in 19 dicotyledonous families are susceptible (Holmes, 1946; Schmelzer, 1967; Edwardson, 1974b; Weinbaum & Milbrath, 1976).
- Diagnostic species
- Nicotiana tabacum
(tobacco). Inoculated leaves usually develop indefinite chlorotic spots or necrotic rings or arcs. Systemic symptoms are vein clearing and necrotic etching, (e.g. Shepherd & Purcifull, 1971), followed by chlorotic mottle (Fig. 1). Necrotic spots and distortion may occur with virulent strains. Numerous other Nicotiana spp. are susceptible. - Datura stramonium. Systemic mottle, leaf distortion (Fig. 2) and vein banding occur. Some
strains reduce the leaves to straps of tissue (Zitter, 1971). Spines on the capsules may be reduced
or absent. This species is not infected by potato virus Y (de Bokx & Huttinga, 1981).
- Capsicum annuum (sweet pepper). Systemic mottle, dark green mosaic and leaf distortion.
- C. frutescens (Tabasco pepper). Most strains induce root necrosis, wilting and death (Greenleaf, 1953).
- Cassia obtusifolia. Necrotic local lesions; distortion, chlorosis, necrosis and stunting of systemically infected leaves.
- Capsicum annuum (sweet pepper). Systemic mottle, dark green mosaic and leaf distortion.
- Propagation species
- Cultivars of tobacco (e.g. Havana 425 and Burley 21) hypersensitive to tobacco mosaic
virus are suitable for maintaining cultures of tobacco etch virus and are good sources of virus and
cytoplasmic (pinwheel) inclusions for purification. D. stramonium is useful as a source for
purification of nuclear inclusions (Dougherty & Hiebert, 1980b).
- Assay species
- The virus induces local lesions in Chenopodium album, C. amaranticolor, C. quinoa and Physalis peruviana. Nicotiana tabacum and Capsicum annuum are useful hosts for aphid transmission studies.
Strains
Variants differing in virulence towards tobacco and other hosts are common. Variants called ‘severe etch’ induce more prominent stunting, chlorosis and necrosis than ‘mild etch’ strains in tobacco (Johnson, 1930; Bawden & Kassanis, 1941). Tobacco cv. V20 is immune to some strains (Christie, Purcifull & Dean, 1974). Other variants include a strain that is not aphid-transmissible (Simons, 1976) and strains that differ in the morphology of the nuclear inclusions they induce in their hosts (Christie & Edwardson, 1977; Edwardson & Christie, 1979).
In SDS-immunodiffusion tests with American Type Culture Collection isolate PV-69 of tobacco etch virus and numerous symptom variants and field isolates of the virus, no antigenic variants were detected (D. Purcifull, unpublished data).
Transmission by Vectors
Transmissible in a non-persistent manner by over 10 species of aphid (Kennedy, Day & Eastop, 1962; Edwardson, 1974a), including Myzus persicae, Macrosiphum euphorbiae and Aphis fabae (Kassanis, 1941). Acquisition and inoculation probes of 10 sec each are sufficient for transmission (Taylor & Robertson, 1974). The virus is retained for only 1-4 h by feeding aphids (Kassanis, 1941) and individual aphids transmit to as many as five consecutive plants (Taylor & Robertson, 1974). Third and fourth instars, as well as adults, can transmit. Pre-access fasting markedly increases transmission. Virus-like particles were observed in the distal portion of the maxillary food canals of viruliferous aphids (Taylor & Robertson, 1974).
A strain of tobacco etch virus that is not normally aphid-transmissible was acquired and transmitted by aphids (M. persicae) that had previously been allowed to probe potato virus Y-infected plants for 1-3 min (Simons, 1976), suggesting, by analogy with potato virus Y (Govier & Kassanis, 1974), that a helper factor is involved. Purified tobacco etch virus was transmissible by aphids only when a helper factor from potato virus Y-infected plants was added to the preparations (Pirone, 1979).
Transmission through Seed
None reported.
Transmission by Dodder
Transmitted by Cuscuta californica, but not by either C. subinclusa or C. campestris (Bennett, 1944). Also transmitted by C. lupuliformis (Schmelzer, 1956).
Serology
Strongly immunogenic. Liquid precipitin tests with plant sap or purified virus have been used for detection and for studying relationships (Chester, 1937; Bartels, 1964; Purcifull, 1964). Untreated virus particles do not diffuse readily in agar gels, but good results are obtained in immunodiffusion tests with particles degraded into low M. Wt diffusible antigens by ethanolamine (Purcifull & Gooding, 1970), acetic acid (Purcifull, 1964), pyrollidine (Shepard, Secor & Purcifull, 1974), or sodium dodecyl sulphate (SDS) (Gooding & Bing, 1970; Purcifull & Batchelor, 1977). For these immunodiffusion tests, antisera prepared to the dissociated proteins are most suitable, though some antisera to untreated virus also react well (Gooding & Bing, 1970). Sonication also fragments the virus into diffusible antigens (H. T. Hsu, unpublished data). Recently the virus has been detected by more sensitive tests, including enzyme-linked immunosorbent assay and passive haemagglutination (H. T. Hsu, unpublished data), and immunosorbent electron microscopy (S. Christie & D. Purcifull, unpublished data). The condition of the coat protein of virus used for serology should be monitored during purification and storage of the virus, because degradation of the protein may occur (E. Hiebert, unpublished data), resulting in a loss of antigenic determinants (Purcifull & Batchelor, 1977). Freeze-dried antigens are reactive and are conveniently stored for use in studies on virus relationships and identification (Purcifull, Christie & Batchelor, 1975).
The cylindrical, cytoplasmic inclusion proteins (Hiebert et al., 1971; Purcifull, Hiebert & McDonald, 1973; Purcifull & Batchelor, 1977) and nuclear inclusion proteins (Knuhtsen, Hiebert & Purcifull, 1974) are immunogenic and detectable by SDS-immunodiffusion procedures (Fig. 3, Fig. 4); they are serologically distinct from each other and from the virus particle protein (Fig. 3).
Immunoprecipitation, using antisera to the various virus-specific antigens, is a useful procedure for analysing in vitro translation products (Dougherty & Hiebert, 1980a, 1980b and 1980c).
Relationships
The chemical and physical properties of the particles, aphid transmissibility, serological relationships and ability to induce pinwheel inclusions in host cells, suggest that tobacco etch virus is a member of the potyvirus group (Matthews, 1979; Hollings & Brunt, 1981). Edwardson (1974a) placed it in Subdivision II of his potyvirus classification system based on the morphology of the pinwheel inclusions and associated lamellae.
Tobacco etch virus is serologically related to, but distinguishable from, several other potyviruses, including potato virus Y (Bartels, 1964; Purcifull, 1964; Purcifull & Gooding, 1970). Shepard, Secor & Purcifull (1974) reported that tobacco etch virus and 14 other potyviruses cross-reacted in immunodiffusion tests with pyrollidine-treated antigens, but in all cases tobacco etch virus was distinguishable from the other potyviruses by spur formation tests. Phenotypic mixing was reported in mixed infections of tobacco etch and potato Y viruses (Nelson & Wheeler, 1981), by immunoelectron microscopy of virus particles in plant extracts. The pinwheel inclusion proteins induced by pepper mottle, potato Y and tobacco etch viruses are serologically related but distinguishable (Purcifull et al., 1973).
The genetic map of tobacco etch virus (see Genome Properties) is similar to those of ten other potyviruses, as deduced by analyses of in vitro translation products (Dougherty & Hiebert, 1980c; E. Hiebert, unpublished data). Antiserum to the larger (54,000 M. Wt) protein subunit of the tobacco etch virus nuclear inclusion reacts with certain in vitro translation products of all 11 potyviruses tested thus far (E. Hiebert, unpublished data).
In cross protection tests, strains protect against one another and may or may not protect against those viruses that are distantly serologically related (Bawden & Kassanis, 1945; Schmelzer, Bartels & Klinkowski, 1960; Laird & Dickson, 1963).
Stability in Sap
In tobacco sap, the thermal inactivation point (10 min) is about 55°C, dilution end-point about 10-4. Infectivity of sap survives for 5-10 days at 20°C. Some infectivity survives for years in dried leaf tissue (McKinney, Silber & Greeley, 1965).
Purification
Dougherty & Hiebert (1980a). Homogenize tobacco tissue (100 g), 3-8 weeks after inoculation, in 150 ml 20 mM Hepes (pH 7.5) containing 0.1% sodium sulphite, and 21 ml n-butanol. Filter and centrifuge at 3300 g for 10 min. To the supernatant fluid add Triton X-100 to 1% (v/v), polyethylene glycol (PEG, M. Wt 6000) to 4% (w/v) and NaCl to 100 mM, and stir for 1 h at 4°C. Centrifuge at 10,000 g for 10 min. Resuspend pellets in 50 ml of 20 mM Hepes (pH 7.5) with the aid of a glass tissue grinder. Centrifuge at 10,000 g for 10 min and discard pellet. Re-precipitate virus from the supernatant fluid by adding PEG to 8% (w/v) and NaCl to 100 mM and centrifuge at 10,000 g for 15 min. Resuspend pellets in 5-10 ml of 20 mM Hepes buffer with the aid of a glass tissue grinder. Layer the resuspended material onto 30% CsCl (w/w) in 20 mM Hepes (pH 7.5) and centrifuge at 140,000 g for 16-18 h at 5°C. Collect virus zone by droplet fractionation, dilute with an equal volume of buffer and then centrifuge at 12,000 g for 10 min. Recover the virus from the supernatant fluid by PEG precipitation as before. Virus yield is about 5-10 mg/100 g of tissue.
Other methods for the purification of tobacco etch virus have been reported (Purcifull, 1966; Damirdagh & Shepherd, 1970a; Hiebert & McDonald, 1973). The use of phosphate buffer during extraction may result in host nucleic acid contamination of the final virus preparation (Dougherty & Hiebert, 1980a). The virus particle protein is liable to undergo limited proteolysis during purification and during storage after purification, presumably due to contaminating and/or adhering host proteases (E. Hiebert, unpublished data).
To purify the cylindrical (pinwheel) inclusions (Dougherty & Hiebert, 1980b), homogenize systemically infected tobacco tissue (5 to 8 weeks after inoculation) in 2 vol. of 0.5 M potassium phosphate buffer (PB), pH 8.2, 0.5% sodium sulphite, and 1 vol. of a 1:1 mixture of chloroform and carbon tetrachloride. The emulsion is separated by centrifugation at 1020 g for 5 min. Re-extract the pellet with 0.5 M PB and centrifuge again. Combine the supernatant fluids and centrifuge at 13,000 g for 15 min. Resuspend the pellets in 0.05 M PB plus 0.1% mercaptoethanol. Add Triton X-100 to 5% (v/v), stir for 1 h and centrifuge the mixture at 27,000 g for 15 min. Resuspend the pellet and layer on a sucrose step gradient (50, 60 and 80%, w/v). After centrifugation at 21,000 rev/min (70,000 g) in a Beckman SW 25.1 rotor for 1 h, recover the inclusions which collect on top of the 80% sucrose zone. Dilute this fraction 4-fold and centrifuge at 27,000 g for 15 min to sediment the inclusions. Yields of up to 25 A280 units (1 A280 unit = 1 mg protein) per 100 g tissue are obtained.
The following method is used to purify nuclear inclusions (Dougherty & Hiebert, 1980b). Homogenize D. stramonium leaf tissue (3 to 4 weeks after inoculation) in 3 vol of 0.1 M PB, pH 7.5, containing 0.2% sodium sulphite. Strain the homogenate through cheesecloth and Miracloth. Add Triton X-100 to 5% (v/v) and stir for 1 h. Centrifuge at 1000 g for 10 min. Resuspend the pellets in 0.02 M PB, pH 8.2, containing 0.5% sodium sulphite. Centrifuge the material again at 1000 g for 10 mm, and resuspend the pellets in 0.02 M PB containing 0.5% sodium sulphite and 40% (w/v) sucrose. Homogenize the material in a Sorvall Omnimixer and then layer onto a 50, 60, 80% (w/v) sucrose discontinuous gradient. Centrifuge the gradients in a Beckman SW 25.1 rotor for 20 min at 15,000 rev/min (40,000 g). Collect the entire 80% sucrose zone and dilute it with 0.02 M PB containing 0.5% sodium sulphite. Sediment the inclusions (1000 g for 10 min) and resuspend them in 0.02 M PB containing 0.5% sodium sulphite and 5% Triton X-100. After stirring for 1 h, sediment the inclusions by centrifugation at 1000 g for 10 min. Resuspend the material in 40% sucrose and centrifuge in sucrose gradients as before. Collect the material that layers on top of the 80% sucrose zone, dilute it with 3 vol of buffer and recover the inclusions by centrifugation. Yields of up to 27 A280 units of protein per 100 g tissue are obtained. Sodium sulphite is required at each step of the purification to preserve the fine substructure of the nuclear inclusions (Fig. 9) (Knuhtsen et al., 1974).
Properties of Particles
Sedimentation coefficient s°20, w: 154 S. No accessory particles are detected by analytical ultracentrifugation (Purcifull, 1966).
A260/A280: 1.15 (uncorrected for light-scattering). Higher values may
indicate contamination with host nucleic acids (Dougherty & Hiebert, 1980a).
Buoyant density: 1.33 g/cm3 (in CsCl).
Extinction coefficient (A0.1%, 1 cm) at 260 nm: 2.4 (Purcifull, 1966).
Particle Structure
Particles are flexuous filaments (Fig. 7, Fig. 8) 12-13 nm in diameter and 730 nm long (Brandes & Wetter, 1959). Leaf dips, examined by immunosorbent electron microscopy, did not reveal the existence of any classes of shorter virus particles that could contain subgenomic RNA molecules (Dougherty & Hiebert, 1980a).
Particle Composition
Nucleic acid: RNA, non-segmented, single-stranded. Molar percentages of nucleotides: G23; A30; C20; U27. RNA is 5% of the particle weight (Damirdagh & Shepherd, 1970b). M. Wt about 3.2 x 106 and sedimentation rate in sucrose gradients about 40 S (Hill & Benner, 1976).
Protein: 95% of particle weight (Damirdagh & Shepherd, 1970b). Subunit M. Wt 3.0-3.2 x l04, but in preparations that have undergone limited proteolysis, for example during storage, the protein M. Wt is only 2.6 x 104 and the amino acid composition is significantly altered (Hiebert & McDonald, 1973; Hiebert et al., 1979). The amino acid composition reported by Damirdagh & Shepherd (1970b) needs to be readjusted to give a subunit M. Wt of 3.0-3.2 x 104.
Genome Properties
Tobacco etch virus RNA has a 3' terminal poly(A) sequence, ranging from about 200 to less than 33 residues but with two size classes, averaging about 150 and 30 residues (Hari, 1981). A genome-linked protein, estimated M. Wt 6000, has been detected but apparently is not required for infectivity (Hari, 1981). Tobacco etch virus RNA is translated efficiently in vitro in rabbit reticulocyte lysates (Dougherty & Hiebert, 1980a, 1980b, 1980c). Four of the six discrete translation products were similar to the four virus-specific proteins found in vivo (virus particle protein, cytoplasmic inclusion protein, and two nuclear inclusion proteins). Two additional discrete products identified in the translation studies have not yet been demonstrated in vivo. Translation products presumed to be readthrough products of adjacent genes on the basis of size, serological reactions and peptide mapping were useful in constructing a genetic map (Fig. 10) (Dougherty & Hiebert, 1980c; E. Hiebert, unpublished data). The proposed genetic map accounts for 95% of the estimated coding capacity of the RNA. The genome-linked protein gene has not been mapped in the tobacco etch virus genome and may represent the remaining 5% of coding capacity.
Tobacco etch virus RNA has also been translated in vitro using the wheat germ system (Koziel et al., 1980; E. Hiebert, unpublished data). The primary product induced in this translation system has a M. Wt of about 3.4 x 104 and does not react with the antisera to the four tobacco etch virus-specific proteins found in vivo (E. Hiebert, unpublished data). This product may represent a premature termination of the gene nearest the 5': end.
Relations with Cells and Tissues
Virus-like particles were observed in plasmodesmata (Weintraub, Ragetli & Leung, 1976) and in the cytoplasm of affected plants (Edwardson, Purcifull & Christie, 1968). Two distinct types of inclusion body also occur in tobacco etch virus-infected cells: crystalline nuclear inclusions and cytoplasmic (pinwheel) inclusions (Kassanis, 1939; Sheffield, 1941; Edwardson, 1974a) (Fig. 5, Fig. 6, Fig. 9).
The nuclear inclusions induced by most strains are truncated pyramids, which appear as squares (Fig. 6) or trapezoids, depending on the plane in which they are observed (Matsui & Yamaguchi, 1964). Other types of nuclear inclusion are associated with specific strains; one is a bi-pyramid (Christie & Edwardson, 1977), and the other is octahedral (Edwardson & Christie, 1979). Nuclear inclusions are readily detected by light microscopy in infected hosts of various plant families (Kassanis, 1939; Purcifull & Edwardson, 1968). Isolated nuclear inclusions show a distinctive cross-hatched substructure (Fig. 9) (Knuhtsen et al., 1974). Subunits in rectangular array were observed in situ (McDonald & Hiebert, 1974a). The nuclear inclusions consist of equimolar amounts of two proteins, of M. Wt 4.9 x 104 and 5.4 x 104. The nuclear inclusion proteins are immunochemically distinct from the virus and from the cytoplasmic inclusions (Knuhtsen et al., 1974; Shepard, Gaard & Purcifull, 1974; Batchelor, 1974).
Cytoplasmic inclusions are composed primarily of pinwheel and laminated aggregate structures (Fig. 5). These inclusions contain a single type of protein of M. Wt 7.0 x 104 (Hiebert & McDonald, 1973). The pinwheel inclusions are serologically distinct from the virus particle protein (Fig. 3), host proteins and nuclear inclusions (Fig. 3) (Shepard & Shalla, 1969; Hiebert et al., 1971; Purcifull et al., 1973; Knuhtsen et al. , 1974). The three-dimensional structures of the pinwheel inclusions associated with tobacco etch virus have been variously interpreted to be cylinders (Edwardson, 1966; Edwardson, Purcifull & Christie, 1968), cones (Andrews & Shalla, 1974), or elliptic hyperboloids (Mernaugh, Gardner & Yocom, 1980). In extracts, the laminated aggregates appear as triangles which have a fine structure consisting of parallel lines with a spacing of 5 nm (Edwardson et al., 1968; Purcifull, Edwardson & Christie, 1970; Shepherd & Purcifull, 1971). Inclusion subunits were observed in freeze-etching studies (McDonald & Hiebert, 1974b).
Notes
Several other members of the potyvirus group, such as potato virus Y ( de Bokx & Huttinga, 1981), pepper mottle virus (Purcifull, Zitter & Hiebert, 1975) and tobacco vein-mottling virus (Sun et al., 1974) occur naturally in solanaceous plants, often in mixed infections with tobacco etch virus. The latter usually can be quickly identified by serological tests (Gooding & Bing, 1970; Purcifull & Batchelor, 1977) and by the characteristic nuclear inclusions found in epidermal strips of its hosts (Kassanis, 1939; Purcifull & Edwardson, 1968; Christie & Edwardson, 1977). Characteristic reactions of necrotic etching in tobacco and wilting of tabasco pepper, the presence of striated, triangular inclusions in leaf extracts, and serological tests with nuclear and cytoplasmic inclusions also are helpful in diagnosis of the virus.
Acknowledgements
Our work on tobacco etch virus was supported by Grants GB-32093, BMS-75-14014, and PCM-7825524 from the National Science Foundation.
Figures

Mottle in systemically infected tobacco leaf.

Distortion and mottle in systemically infected Datura stramonium leaf.
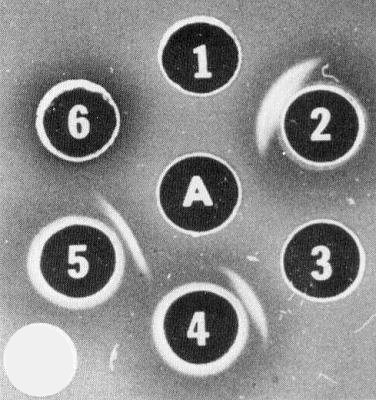
Serological unrelatedness of particle protein, nuclear inclusion proteins and cytoplasmic (pinwheel) inclusion protein of tobacco etch virus. A = purified nuclear inclusions; 1 = purified virus; 2 = virus antiserum; 3 = purified cytoplasmic inclusions; 4 = cytoplasmic inclusion antiserum; 5 = nuclear inclusion antiserum; 6 = concentrated extracts from healthy Datura plants. Agar medium contains 0.5% sodium dodecyl sulphate. (Reprinted from Knuhtsen et al. (1974) by permission of Academic Press, Inc.)

Detection of tobacco etch virus and inclusion antigens in sodium dodecyl sulphate (SDS) treated plant extracts by the immunodiffusion test. A = extract from tobacco etch virus-infected D. stramonium; 1 = antiserum to SDS-treated nuclear inclusions; 2 and 6 = extract from healthy Datura; 3 = antiserum to SDS-treated cytoplasmic (pinwheel) inclusion; 4 = normal serum; 5 = antiserum to SDS-treated virus.

Electron micrograph of pinwheel and laminated aggregate inclusions in cytoplasm of infected tobacco leaf cell (courtesy of J. R. Edwardson and R. G. Christie). Bar represents 500 nm.
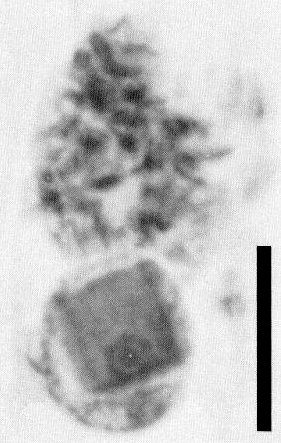
Light micrograph of infected tobacco epidermal cell, showing square inclusion in nucleus and massive cytoplasmic inclusion above (courtesy of R. G. Christie and J. R. Edwardson). Bar represents 10 µm.
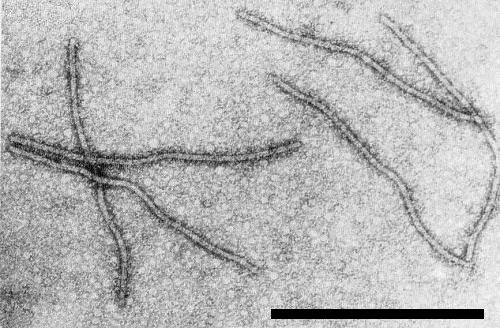
Electron micrograph of tobacco etch virus particles in leaf extract stained with uranyl formate (courtesy of S. R. Christie). Bar represents 500 nm.

Substructure of a virus particle in electron micrograph of extract mounted in uranyl formate (courtesy of S. R. Christie).

Electron micrograph of a portion of a tobacco etch virus-induced nuclear inclusion, stained in uranyl acetate. Bar represents 50 nm.
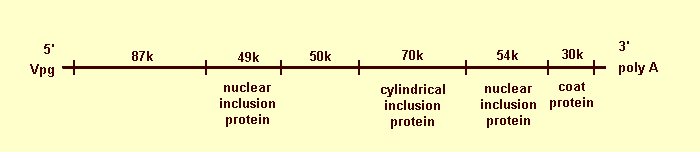
Proposed genetic map for tobacco etch virus. The M. Wts of the gene products are presented above the map, and four of the products are identified below the map. A genome-linked protein (Vpg) is indicated at the 5' end, and polyadenylate (poly A) at the 3' end.
References list for DPV: Tobacco etch virus (258)
- Anderson, Pl. Dis. Reptr 38: 736, 1954.
- Anderson, Phytopathology 49: 97, 1959.
- Andrews & Shalla, Phytopathology 64: 1234, 1974.
- Bartels, Phytopath. Z. 49: 257, 1964.
- Batchelor, Ph.D. Diss., Univ. Fla, 82 pp., 1974.
- Bawden & Kassanis, Ann. appl. Biol. 28: 107, 1941.
- Bawden & Kassanis, Ann. appl. Biol. 32: 52, 1945.
- Bennett, Phytopathology 34: 905, 1944.
- Blakeslee, J. Genet. 11: 17, 1921.
- Brandes & Wetter, Virology 8: 99, 1959.
- Chester, Phytopathology 27: 903, 1937.
- Christie & Edwardson, Monograph Ser. Fla agric. Exp. Stn 9, 150 pp., 1977.
- Christie, Purcifull & Dean, Pl. Dis. Reptr 58: 658, 1974.
- Cook, Ozaki, Zitter & Blazquez, Circ. S-242, Univ. Fla agric. Exp. Stn 6 pp., 1976.
- Cook, Zitter & Ozaki, Circ. S-251, Univ. Fla agric. Exp. Stn, 5 pp., 1977.
- Damirdagh & Shepherd, Phytopathology 60: 132, 1970a.
- Damirdagh & Shepherd, Virology 40: 84, 1970b.
- de Bokx & Huttinga, CMI/AAB Descr. Pl. Viruses 242, 6 pp., 1981.
- Debrot, Agronomia trop. 26: 321, 1976.
- Demski, Pl. Dis. Reptr 63: 647, 1979.
- Dougherty & Hiebert, Virology 101: 466, 1980a.
- Dougherty & Hiebert, Virology 104: 174, 1980b.
- Dougherty & Hiebert, Virology 104: 183, 1980c.
- Edwardson, Science N.Y. 153: 883, 1966.
- Edwardson, Monograph Ser. Fla agric. Exp. Stn 4, 398 pp., 1974a.
- Edwardson, Monograph Ser. Fla agric. Exp. Stn 5, 225 pp., 1974b.
- Edwardson & Christie, Fitopatol. Brasileira 4: 341, 1979.
- Edwardson, Purcifull & Christie, Virology 34: 250, 1968.
- Gooding, Pl. Dis. Reptr 54: 119, 1970.
- Gooding & Bing, Phytopathology 60: 1293, 1970.
- Gooding & Ross, Tob. Sci. 14: 55, 1970.
- Govier & Kassanis, Virology 61: 420, 1974.
- Greenleaf, Phytopathology 43: 564, 1953.
- Hari, Virology 112: 391, 1981.
- Hiebert & McDonald, Virology 56: 349, 1973.
- Hiebert, Purcifull, Christie & Christie, Virology 43: 638, 1971.
- Hiebert, Tremaine & Ronald, Phytopathology 69: 1031, 1979.
- Hill & Benner, Virology 75: 419, 1976.
- Hollings & Brunt, CMI/AAB Descr. Pl. Viruses 245, 7 pp., 1981.
- Holmes, Phytopathology 36: 643, 1946.
- Johnson, Bull. Ky agric. Exp. Stn 306: 289, 1930.
- Kassanis, Ann. appl. Biol. 26: 705, 1939.
- Kassanis, Ann. appl. Biol. 28: 238, 1941.
- Kennedy, Day & Eastop, A conspectus of aphids as vectors of plant viruses, London, Commonwealth Institute of Entomology, 114 pp., 1962.
- Knuhtsen, Hiebert & Purcifull, Virology 61: 200, 1974.
- Koziel, Hari & Siegel, Virology 106: 177, 1980.
- Laird & Dickson, Phytopathology 53: 48, 1963.
- Matsui & Yamaguchi, Virology 22: 40, 1964.
- Matthews, Intervirology 12: 258, 1979.
- McDonald & Hiebert, J. Ultrastruct. Res. 48: 138, 1974a.
- McDonald & Hiebert, Virology 58: 200, 1974b.
- McKinney, Silber & Greeley, Phytopathology 55: 1043, 1965.
- Mernaugh, Gardner & Yocom, Virology 106: 273, 1980.
- Nelson & Wheeler, Phytopathology 71: 245, 1981.
- Pirone, Phytopathology 69: 531, 1979.
- Purcifull, Ph.D. Diss., Univ. Calif. Davis, 80 pp., 1964.
- Purcifull, Virology 29: 8, 1966.
- Purcifull & Batchelor, Bull. Fla agric. Exp. Stn 788, 39 pp., 1977.
- Purcifull & Edwardson, Phytopathology 58: 532, 1968.
- Purcifull & Gooding, Phytopathology 60: 1036, 1970.
- Purcifull, Edwardson & Christie, Phytopathology 60: 779, 1970.
- Purcifull, Hiebert & McDonald, Virology 55: 275, 1973.
- Purcifull, Zitter & Hiebert, Phytopathology 65: 559, 1975.
- Purcifull, Christie & Batchelor, Phytopathology 65: 1202, 1975.
- Schmelzer, Phytopath. Z. 28: 1, 1956.
- Schmelzer, Phytopath. Z. 60: 301, 1967.
- Schmelzer, Bartels & Klinkowski, Phytopath. Z 40: 52, 1960.
- Sheffield, Jl R. microsc. Soc. 61: 30, 1941.
- Shepard & Shalla, Virology 38: 185, 1969.
- Shepard, Gaard & Purcifull, Phytopathology 64: 418, 1974.
- Shepard, Secor & Purcifull, Virology 58: 464, 1974.
- Shepherd & Purcifull, CMI/AAB Descr. Pl. Viruses 55, 4 pp., 1971.
- Simons, Phytopathology 66: 652, 1976.
- Stover, Phytopathology 41: 1125, 1951.
- Sun, Gooding, Pirone & Tolin, Phytopathology 64: 1133, 1974.
- Taylor & Robertson, Phytopath. Z. 80: 257, 1974.
- Weinbaum & Milbrath, Pl. Dis. Reptr 60: 469, 1976.
- Weintraub, Ragetli & Leung, J. Ultrastruct. Res. 56: 351, 1976.
- Zitter, Proc. Fla St. hort. Soc. 84: 177, 1971.