Details of DPV and References
DPV NO: 38 October 1970
Family: Secoviridae
Genus: Nepovirus
Species: Tomato black ring virus | Acronym: TBRV
Tomato black ring virus
A. F. Murant Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Smith (1946).
- Selected synonyms
- Buschbohnenringflecken-Virus = bean ringspot virus (Rev. appl. Mycol.
35: 411; 42: 457)
- Lettuce ringspot virus (Rev. appl. Mycol. 39: 142)
- Kartoffelbukett-Virus = potato bouquet virus (Rev. appl. Mycol. 32: 274; 37: 463)
- (For names of other more serologically distinctive isolates see also under Strains).
- Lettuce ringspot virus (Rev. appl. Mycol. 39: 142)
- A RNA-containing virus which has isometric particles about 30 nm in diameter and occurs in Europe. It is readily sap-transmissible, has a wide host range, infects the seed of many host plants, and is transmitted in soils by nematodes (Longidorus spp.).
Main Diseases
In addition to black ring of tomato, the various strains of this virus cause ringspot diseases of bean, sugar beet, lettuce, raspberry and strawberry, yellow vein of celery, ‘bouquet’ and pseudo-aucuba of potato, shoot-stunting of peach, one type of mosaic of Robinia pseudoacacia and unnamed diseases of leek and onion. It occurs in many other plants, including cabbage, grapevine, lucerne and Sambucus nigra.
Geographical Distribution
Not reported outside Europe.
Host Range and Symptomatology
Host range is wide: infects naturally many species of wild and cultivated monocotyledonous and dicotyledonous plants. An isolate from Robinia pseudoacacia infected 76 species in 29 dicotyledonous families after mechanical inoculation (Schmelzer, 1963). It is reported to infect the roots of the gymnosperm, Picea sitchensis. Nearly all commonly used herbaceous test plants are susceptible.
- Diagnostic species
- Chenopodium amaranticolor and C. quinoa. Chlorotic or necrotic
local lesions (Fig. 1); systemic necrosis or chlorotic mottle (Fig. 2).
- Nicotiana rustica and N. tabacum cvs. White Burley or Xanthi-nc (tobacco). Local chlorotic or necrotic spots or rings (Fig. 3); systemic spots, rings and line patterns with variable amounts of necrosis (Fig. 4). Leaves produced later appear normal but contain virus.
- Phaseolus vulgaris cv. The Prince (French bean). In winter in the U.K., dark brown local lesions c. 2 mm in diameter (Fig. 5), in summer chlorotic lesions or none; systemic chlorotic mottle with variable amounts of necrosis and distortion.
- Petunia hybrida. Chlorotic local lesions sometimes with brown necrotic margins; systemic veinal chlorosis or necrosis. Leaves produced subsequently are symptomless but contain virus.
- Nicotiana rustica and N. tabacum cvs. White Burley or Xanthi-nc (tobacco). Local chlorotic or necrotic spots or rings (Fig. 3); systemic spots, rings and line patterns with variable amounts of necrosis (Fig. 4). Leaves produced later appear normal but contain virus.
- Propagation species
- Nicotiana rustica is a suitable plant for maintaining cultures; N. clevelandii and
Petunia hybrida are good sources of virus for purification.
- Assay species
- Chenopodium amaranticolor is the most reliable local-lesion host. C. quinoa, Cucumis sativus (cucumber) and Brassica rapa (turnip) are convenient ‘bait plants’ in nematode transmission experiments.
Strains
The best known strains are:
Tomato black ring strain (Smith, 1946). The type strain.
Lettuce ringspot strain (Smith & Short, 1959).
Potato bouquet strain of Köhler, Rev. appl. Mycol. 32: 274 (Harrison, 1958).
Potato pseudo -aucuba strain of Köhler, Rev. appl. Mycol. 35: 481 (Bercks, 1962).
Beet ringspot strain (Harrison, 1957).
Celery yellow vein strain (Hollings, 1965).
These strains probably cannot be reliably distinguished by symptoms on host plants but differ in their serological and vector relationships.
Transmission by Vectors
Vectors are free-living soil-inhabiting nematodes of the genus Longidorus. Strains serologically similar to the type strain are transmitted most efficiently by L. attenuatus, those more closely related to the beet ringspot strain by L. elongatus (Harrison, Mowat & Taylor, 1961; Harrison, 1964; Taylor & Murant, 1969). Larvae and adults of L. elongatus both transmit, but the adult does not pass the virus to its progeny, nor is the virus retained when the nematode moults. L. elongatus kept in fallow soil retains infectivity up to about 9 weeks. Virus-like particles seem adsorbed to the stylet guiding sheath of L. elongatus which have fed on plants infected with tomato black ring virus (Taylor & Robertson, 1969).
Transmission through Seed
Reported in at least twenty-four spp. in fifteen botanical families. In many hosts more than 10%, and in some 100%, of progeny seedlings are infected. Many plants infected through the seed show no symptoms. Virus was transmitted to seed of raspberry and strawberry from either male or female parent, but plants pollinated with virus-carrying pollen did not become infected (Lister & Murant, 1967). Besides aiding dissemination of the virus, infection of the seed seems to provide an important means of survival of the virus in soils (Murant & Lister, 1967).
Transmission by Dodder
Nine species of Cuscuta failed to transmit the potato bouquet strain (Schmelzer, 1956).
Serology
Strongly immunogenic in rabbits; antisera with titres of 1/500 are easily obtained. Precipitin tests in tubes, droplets or 1% agar gel give satisfactory results. The virus gives one line of precipitate in gel-diffusion tests. The celery yellow vein strain is unusual in eliciting no antibodies in rabbits unless virus treated with 0.2% formaldehyde is used as immunogen (Hollings & Stone, 1962).
Relationships
There is much antigenic variation between strains. Most antisera place the strains into two groups; one contains the type, lettuce ringspot and potato bouquet strains; the other contains the beet ringspot and potato pseudo-aucuba strains (Harrison, 1958; Bercks, 1962); the celery yellow vein strain seems intermediate (Hollings, 1965).
Plant-protection may not be complete, even between strains that are closely serologically related, and is frequently only partial or non-reciprocal in tests between distantly related strains.
Tomato black ring virus shares many properties with other nepoviruses (arabis mosaic, grapevine fanleaf, raspberry ringspot, strawberry latent ringspot, tobacco ringspot and tomato ringspot) but is unrelated to them serologically.
Stability in Sap
In tobacco sap, the virus usually loses infectivity after 10 min at 60-65°C storage at room temperature for 2-3 weeks, or dilution to 10-3-10-4. The lesion number decreases with dilution by more than the dilution factor (Harrison, 1957).
Purification
A modification of Steere’s butanol/chloroform method is useful (Harrison & Nixon, 1960).
Properties of Particles
The particles are all the same size but sediment as three components (Fig. 6), empty protein shells (T) and two kinds of nucleoprotein with different amounts of RNA (M and B).
Sedimentation coefficients (s20,w) at infinite dilution (svedbergs): 55(T), 97(M), 121(B).
Other physical properties not determined.
Particle Structure
Isometric, about 30 nm in diameter with a 5- or 6-sided angular outline (Harrison & Nixon, 1960). Electron micrographs show particles some completely, some partially and some not penetrated by negative stain (Fig. 7). These particles possibly correspond to the T, M and B components. Detailed structure of the particle not known.
Particle Composition
RNA about 28% (M) and 38% (B) of the particle weight (estimated from the sedimentation coefficients).
Relations with Cells and Tissues
No information.
Notes
Plants infected with tomato black ring virus are patchily distributed in crops because of the slow migration of the soil-inhabiting vectors, Longidorus spp. Some strains tend to occur in soils together with strains of raspberry ringspot virus because they share the vector, L. elongatus. These two viruses are serologically unrelated; they can be distinguished from each other by their reactions in Chenopodium amaranticolor. In general, however, these and other unrelated viruses of the nepovirus group cannot be reliably identified by host range or symptomatology; serological tests are essential.
Figures

Local lesions in Chenopodium amaranticolor.

Systemic necrosis in C. quinoa.
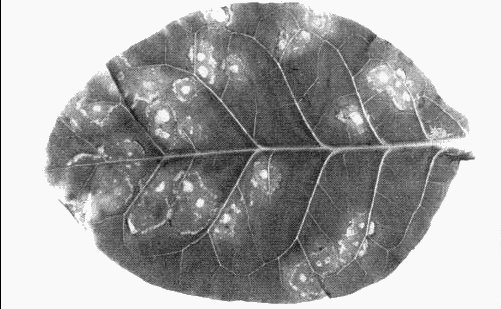
Local necrotic rings and spots in Nicotiana tabacum, cv. White Burley.
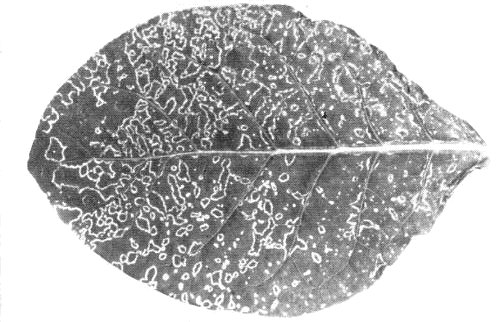
Systemic symptoms in N. tabacum cv. Xanthi-nc.

Local lesions in Phaseolus vulgaris cv. The Prince.
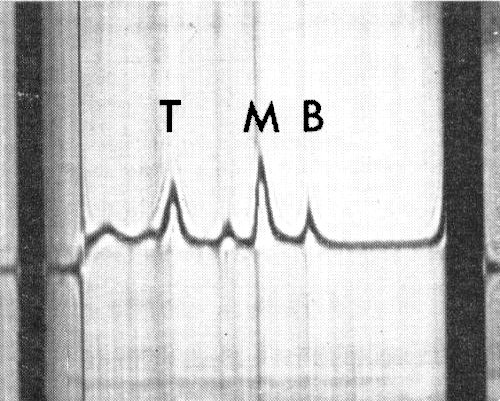
Schlieren diagram of a sedimentation analysis of a partially purified preparation, showing three virus components (T, M and B). The other components were probably derived from the N. clevelandii host tissue.
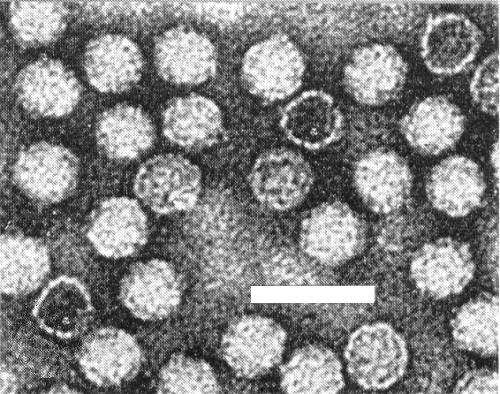
Virus particles from a purified preparation showing some particles completely, some partially and some not penetrated by phosphotungstate. Bar represents 50 nm.
References list for DPV: Tomato black ring virus (38)
- Bercks, Phytopath. Z. 46: 97, 1962.
- Harrison, Ann. appl. Biol. 45: 462, 1957.
- Harrison, J. gen. Microbiol. 18: 450, 1958.
- Harrison, Virology 22: 544, 1964.
- Harrison, Mowat & Taylor, Virology 14: 480, 1961.
- Harrison & Nixon, Virology 12: 104, 1960.
- Hollings, Ann. appl. Biol. 55: 459, 1965.
- Hollings & Stone, Nature, Lond. 194: 607, 1962.
- Lister & Murant, Ann. appl. Biol. 59: 49, 1967.
- Murant & Lister, Ann. appl. Biol. 59: 63, 1967.
- Schmelzer, Phytopath. Z. 28: 1, 1956.
- Smith, Parasitology 37: 126, 1946.
- Smith & Short, Pl. Path. 8: 54, 1959.
- Taylor & Murant, Ann. appl. Biol. 64: 43, 1969.
- Taylor & Robertson, Ann. appl. Biol. 64: 233, 1969.
- Schmelzer, Phytopath. Z. 46: 235, 1963.