Details of DPV and References
DPV NO: 308 September 1985
Family: Tombusviridae
Genus: Dianthovirus
Species: Carnation ringspot virus | Acronym: CRSV
There is a more recent description of this virus: DPV 371
This is a revised version of DPV 21
Carnation ringspot virus
J. H. Tremaine Agriculture Canada Research Station, 6660 NW. Marine Drive, Vancouver, B.C., Canada, V6T 1X2
J. A. Dodds Department of Plant Pathology, University of California, Riverside, California, USA 92521
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Kassanis (1955).
Synonym
- Anjermozaiek virus (Rev. appl. Mycol. 31: 122)
-
A virus with isometric (icosahedral) particles 34 nm in diameter in which two species of single-stranded RNA of messenger polarity are separately packaged. Both RNA species are needed for infectivity. The virus is transmissible by vegetative propagation, by inoculation of sap and by foliage and root contact. It is believed to be soil-borne; nematode transmission has been reported but requires confirmation. Once common wherever carnations are cultivated, but can be controlled by use of virus-tested stock plants. Also detected in fruit tree species.
Main Diseases
Leaf mottle, ringspots, stunting and distortion in carnation and Dianthus barbatus (Sweet William), sometimes with leaf tip necrosis (Fig. 1; Hollings & Stone, 1970; Lovisolo & Lisa, 1978). Flowers distorted and of poor quality (Fig. 2). Associated with a stony pit disease of pears (Richter et al., 1978), and decline diseases of sour cherry and apple (Kleinhempel et al., 1980). Also occurs in the weed Stellaria media (Rudel et al., 1977).
Geographical Distribution
Found wherever carnations are grown by vegetative propagation in temperate regions (Bremer & Lahdenpera, 1981; Lommel et al., 1983). Isolated from apple, pear and sour cherry in the German Democratic Republic (Kleinhempel et al., 1980; Richter et al., 1978).
Host Range and Symptomatology
Over 133 species in 25 dicotyledonous families can be infected experimentally (Hollings & Stone, 1970; Kleinhempel et al., 1980). Transmissible mechanically, but more easily from sap of Nicotiana clevelandii or Vigna unguiculata ssp. sinensis than from carnation.
-
Diagnostic species
- Dianthus barbatus
(Sweet William). Inoculated leaves show flecks, rings and ringspots after 4-7 days followed by systemic chlorotic and semi-necrotic rings and flecks. Not all clones show obvious symptoms or support large concentrations of virus. - Gomphrena globosa. Local necrotic rings develop in 2-4 days after
inoculation followed by systemic flecking, mottle and distortion.
- Phaseolus vulgaris (French bean). Local chlorotic dots in 4-5 days, becoming white and necrotic; irregular systemic spotting and necrotic veinal flecks, later growth symptomless though infected.
- Chenopodium amaranticolor and C. quinoa. Local necrotic lesions in 2-4 days (Fig. 3); usually not systemic.
- Tetragonia expansa. Local white necrotic dots in 2-3 days, sometimes followed by systemic chlorotic flecks.
- Vigna unguiculata ssp. sinensis (cowpea). Local necrotic lesions in 2-4 days (Fig. 4), followed by systemic mottling, necrotic flecking, rugosity and rolling of leaf laminae (Weintraub et al., 1975).
- Phaseolus vulgaris (French bean). Local chlorotic dots in 4-5 days, becoming white and necrotic; irregular systemic spotting and necrotic veinal flecks, later growth symptomless though infected.
-
Propagation species
- Dianthus barbatus
is a suitable host for maintaining cultures. Nicotiana clevelandii. Phaseolus vulgaris and Vigna unguiculata ssp. sinensis are good hosts from which to purify the virus.Assay species
- Chenopodium amaranticolor, C. quinoa
and Vigna unguiculata ssp. sinensis are useful local lesion hosts.
Strains
Strains with differences in host reactions have not been reported. However, Tremaine et al. (1976, 1983, 1984) and Tremaine & Ronald (1976) have described strains (N, A and R) that differ in such properties as particle aggregation forms in purified preparations, effects of sodium dodecyl sulphate (SDS) and sodium dextran sulphate on particle stability, amino acid composition and serological properties (see Properties of Particles).
Transmission by Vectors
Transmission is reported by Longidorus elongatus, L. macrosoma and Xiphinema diversicaudatum (Fritzsche & Schmelzer, 1967; Fritzsche et al., 1979), but the evidence supporting these claims is considered to be inadequate (Trudgill et al., 1983; Brown & Trudgill, 1984). Spread in orchard soils without vector involvement is suspected (Kegler & Kegler, 1981).
Transmission through Seed
None found.
Serology
Very immunogenic. Antisera with tube precipitin titres of 1/4000 or higher and gel diffusion precipitin titres of up to 1/1024 have been obtained. Virus can usually be detected in carnation sap by gel diffusion tests. Both double antibody sandwich ELISA and indirect ELISA (antigen/diluted sap as first reagent) are useful for indexing (Lommel et al., 1982). Antibodies in antisera prepared by injecting rabbits with preparations of virus particles do not react in gel diffusion tests with protein subunits dissociated by EDTA-NaCl or by SDS but do react with these antigens in indirect ELISA (J. H. Tremaine, unpublished data). Strain differences have been detected in gel diffusion tests (Tremaine et al., 1976, 1984).
Relationships
In physical and chemical properties, carnation ringspot virus is very similar to red clover necrotic mosaic virus and the two viruses are placed together in the dianthovirus group (Hollings & Stone, 1977; Matthews, 1982; Morris-Krsinich et al., 1983). These viruses form stable pseudo-recombinants, indicating a close genetic relationship (Lommel & Morris, 1982; Lommel, 1983) and preparations of their particles exhibit weak serological cross-reactions in indirect ELISA (Hiruki et al., 1984). Stronger serological cross-reactions were found when the dissociated virus proteins were compared by the same method (J. H. Tremaine, unpublished data). Unexpected serological cross-reactions with viruses of the tymovirus group have been reported (Bercks & Querfurth, 1972).
Stability in Sap
In sap of Dianthus barbatus, the thermal inactivation point (10 min) is about 80°C although much infectivity is lost above 60°C. Dilution end-point is 10-5 and infectivity is retained at 20°C for 50-60 days. In lyophilized sap of Nicotiana clevelandii the virus retained infectivity at room temperature, under vacuum, for over 6 years (Hollings & Stone, 1970).
Purification
Easily purified from infected cowpea or Nicotiana clevelandii plants. Fresh tissue can yield 150 mg virus/kg; frozen tissue typically yields less.
Method 1 (Tremaine et al., 1976). Extract 100 g infected tissue 3 weeks after inoculation in 200 ml 0.2 M sodium acetate buffer (pH 5.0) containing 0.02 M sodium diethyl dithiocarbamate and 0.1% 2-mercaptoethanol. Adjust the extract to pH 5.0 with 10% acetic acid, and leave at 5°C for 4 h. Clarify by low speed centrifugation and precipitate the virus by adding polyethylene glycol (M. Wt 6000) to 8.0% (w/v). Suspend the pellet from low speed centrifugation in 0.1 M sodium acetate buffer (pH 5.0) and give the preparation one cycle of differential centrifugation (20 min at 12,000 g; 90 min at 95,000 g). Further purify by repeated cycles of differential centrifugation and rate zonal sucrose density gradient centrifugation.
Method 2 (Hollings & Stone, 1965). Must be used for strains that precipitate at pH 5.0. Extract each 100 g tissue in 125 ml 0.05 M phosphate buffer (pH 7.6) containing 0.1 % thioglycollic acid. Clarify by stirring with 8.5% n-butanol overnight at 2°C. The virus is then concentrated and purified by differential centrifugation.
Properties of Particles
The particles of the virus swell above pH 7.0; the swelling can be prevented or reversed by divalent cations. The particles dissociate into RNA and protein in 0.1 M Tris-HCl buffer, pH 7.5, containing 10 mM EDTA and 1 M NaCl, and stable virus particles can be reconstituted by dialysis in 0.1 M Tris-HCl buffer, pH 7.0, containing divalent cations (J. H. Tremaine, unpublished data). Particles are stabilized by pH-dependent protein-protein interactions and by RNA-protein interactions. Particles of the A strain can form aggregates of six virus particles and linked aggregates (Fig. 7, Fig. 8, Fig. 9, Fig. 10, Fig. 11, Fig. 12). Particles of the R and N strains aggregate in a temperature-reversible manner but the transition occurs at lower temperatures with particles of the R strain than with those of the N strain (Fig. 6). Particles of the N strain are readily dissociated at room temperature in low concentrations of SDS or sodium dextran sulphate at pH 7.0. Particles of the A and R strains are more stable than particles of the N strain (Tremaine & Ronald, 1976; Ronald & Tremaine, 1976; Tremaine et al., 1976, 1983, 1984).
Sedimentation coefficient (s20,w): N strain, 133 S at pH 5.0, 125 S at pH 7.5 (J. H. Tremaine, unpublished data). Particle weight (daltons): 7.07 x 106 (Kalmakoff & Tremaine, 1967); 8.3 x 106 calculated assuming 180 protein subunits of 38,000 daltons and RNA-1 of 1.5 x 106 daltons.
Diffusion coefficient (D20,w x 10-7 cm2/sec): 1.48 (Kalmakoff & Tremaine, 1967).
Partial specific volume: 0.693 cm3/g; calculated from percentage nucleic acid, amino acid and nucleotide composition (Kalmakoff & Tremaine, 1967).
Electrophoretic mobility: in free boundary electrophoresis the virus particles are isoelectric at pH 4.5; at pH 6.0 to 8.0 the virus boundary moves to the anode as an increasingly broad schlieren peak which continues to broaden when the polarity is reversed; migration in agarose is similar (J. H. Tremaine, unpublished data).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 6.46 (Kalmakoff & Tremaine, 1967).
A260/A280: 1.67, corrected for light-scattering (J. H. Tremaine, unpublished data).
Buoyant density (g/cm3): 1.366 in CsCl; when limited proteolysis of virus occurs, additional species with buoyant densities of 1.369 to 1.374 are also present (Fig. 5); 1.275 in metrizamide (J. H. Tremaine, unpublished data).
Particle Structure
Particles are isometric, approximately 34 nm in diameter (Fig. 13) and have a T = 3 structure with 180 protein subunits. The particles are stable in phosphotungstate and uranyl acetate.
Particle Composition
Nucleic acid: Single-stranded RNA, about 20% of the particle weight (Kalmakoff & Tremaine, 1967). Two genomic species (Fig. 14) of M. Wt (x 10-6) c. 1.5 (RNA-1) and 0.5 (RNA-2) are released from the particles in similar amounts (Dodds et al., 1977; Lommel, 1983). Particle properties and composition suggest that there are two classes of particle, one containing RNA-1, the other containing more than one (probably three) copies of RNA-2. Molar percentages of nucleotides G26; A27; C23; U24 (Kalmakoff & Tremaine, 1967).
Protein: Coat protein can be isolated by first swelling the virus at 5-10 mg/ml in 0.1 M Tris-HCl, 0.01 M EDTA (pH 7.5 to 8.0) for at least 1 h at 0°C and then adding an equal volume of 2.0 M NaCl and subjecting the mixture to rate zonal density gradient centrifugation in a 5-35% sucrose density gradient in 0.1 M Tris-HCl buffer, pH 7.0, containing 1.0 M NaCl (J. H. Tremaine, unpublished data). Virus particles contain a single polypeptide species comprising approximately 80% of the particle weight. The polypeptide has a M. Wt of 38,000 but limited proteolysis may occur in stored preparations to yield polypeptides of M. Wt 36,000 and 34,000. The polypeptide contains about 347 amino acid residues but the amino acid composition differs for the A, R and N strains (Tremaine et al., 1984).
Genome Properties
Both RNA-1 and RNA-2 are required for infectivity. Determinants for the particle protein and host range are found on RNA-1 (Dodds et al., 1977; Henriques & Morris, 1979; Lommel, 1983). No function has yet been assigned to RNA-2. Translation of unfractionated RNA in the rabbit reticulocyte lysate or wheat germ cell-free systems results in three major products of M. Wt 34,000, 36,000 and 39,000 (Lommel, 1983; J. H. Tremaine, unpublished data). Two double-stranded RNA species twice the weight of, and sharing nucleotide sequences with, single-stranded RNA-1 and RNA-2 respectively, are present in extracts of infected plants (Fig. 14). An additional unexpected dsRNA species with an electrophoretic mobility similar to that of dsRNA-2 is also present (Dodds et al., 1977; Henriques & Morris, 1979), and shares sequence homology with RNA-1, but not RNA-2 (Lommel, 1983). Preparations of dsRNA are not infective but become so on melting (Dodds et al., 1977). The genome organization has features in common with that of pea enation mosaic virus in that the determinant for the coat protein is located on the larger of the two genomic RNA species (Lane, 1979). There is no sequence homology between RNA-1 and RNA-2 and neither RNA possesses a polyadenylated region. Pseudo-recombinants can be constructed between RNA-1 from the N strain and RNA-2 from the A strain, and vice versa (Dodds et al., 1977). Stable pseudo-recombinants have also been formed between carnation ringspot virus RNA-2 and red clover necrotic mosaic virus RNA-1 (Lommel & Morris, 1982; Lommel, 1983). There is no detectable sequence homology between the RNA molecules of carnation ringspot virus and red clover necrotic mosaic virus and the identity of the pseudo-recombinants was established by cDNA hybridization analysis.
Relations with Cells and Tissues
The virus is found in stems, leaves, flowers and roots, and in some apical meristems. Free virus particles occur in the cytoplasm and in the nucleoplasm of infected mesophyll cells of Dianthus barbatus, and virus crystals are found in the cytoplasm of older infected leaves of cowpea (Fig. 15). No other inclusion bodies or sites of synthesis have been reported except for the appearance of abnormal tubular (32 nm diameter) and spherical (60-65 nm diameter) structures in the nuclei of mesophyll cells of infected Dianthus barbatus (Fig. 16) (Weintraub et al., 1975).
Notes
Carnation ringspot virus can be readily eliminated from several hosts by heat treatment and also by meristem-tip culture (Rybalko & Kharuta, 1978). It can be controlled by an eradication programme based on adequate serological indexing (Lommel et al., 1983). It is commonly found in carnation together with carnation mottle virus from which it can be readily distinguished by its symptoms in Gomphrena globosa and Dianthus barbatus. Specific antisera will also distinguish between these two viruses. The symptoms caused by carnation ringspot virus in carnation cannot readily be confused with those caused by any other carnation virus. Carnation Italian ringspot virus is a different virus, belonging to the tombusvirus group (Hollings et al., 1970).
Acknowledgements
We would like to thank S. Lommel for reading the final draft
and providing a figure. We also wish to acknowledge M. Hollings and O. Stone, who
prepared the first CMI/AAB Description of this virus (No. 21). Text and figures
from Descr. No. 21 were used in part to prepare this revised description.
Photographs: 1, 2, 3 and 4, courtesy M. Hollings & O. Stone; 14, courtesy S. Lommel; 15 and 16, courtesy M. Weintraub.
Figures

(Left ) Healthy ‘Joker’ carnation plant, (right) plant infected with carnation ringspot virus.

(Left) Healthy ‘Joker’ carnation flower, (right) flower infected with carnation ringspot virus.

Local lesions in Chenopodium amaranticolor.

Local lesions in Vigna unguiculata ssp. sinensis.
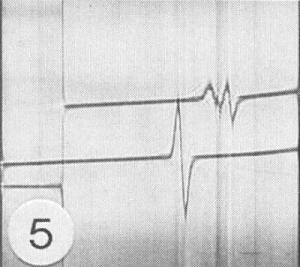
Schlieren patterns of equilibrium sedimentation analyses in CsCl of two preparations of the N strain. (Top) Stored virus preparation with partially proteolysed virus coat protein showing three components with buoyant densities of 1.369, l.37l and l.379 g/cm3. (Bottom) Freshly prepared virus preparation without partial proteolysis showing single component with buoyant density of 1.368 g/cm3. Menisci are at the left.
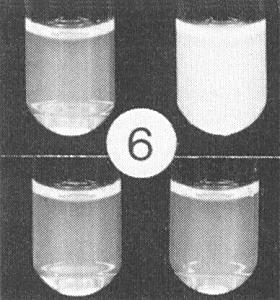
(Top) Two aliquots of preparation of reversibly aggregating strain R, (left) at 4°C, (right) at 30°C, (bottom) same aliquots after cooling to 4°C.
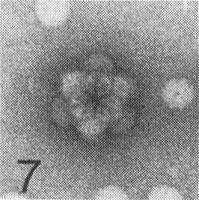
Electron micrograph of particles of strain A negatively stained with 1% uranyl acetate. Cluster of 12 virus particles seen from the three-fold axis. Bar represents 100 nm.

Electron micrograph of particles of strain A negatively stained with 1% uranyl acetate. A similar cluster to that shown in Fig. 7 seen from the five-fold axis. Bar represents 100 nm.

Electron micrograph of particles of strain A negatively stained with 1% uranyl acetate; dimer cluster of 23 virus particles. Bar represents 100 nm.
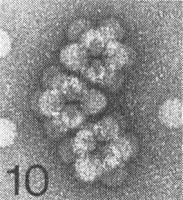
Electron micrograph of particles of strain A negatively stained with 1% uranyl acetate; angled trimer cluster. Bar represents 100 nm.

Electron micrograph of particles of strain A negatively stained with 1% uranyl acetate; linear trimer cluster. Bar represents 100 nm.
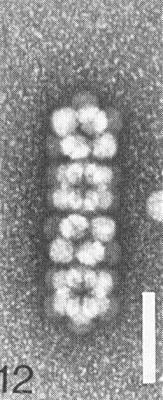
Electron micrograph of particles of strain A negatively stained with 1% uranyl acetate; linear tetramer cluster. Bar represents 100 nm.
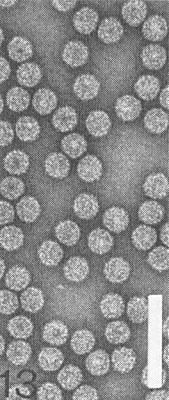
Electron micrograph of virus particles of N strain. Bar represents 100 nm.

(Left) Agarose gel electrophoretic patterns of genomic ssRNA species isolated from virus particles, (right) polyacrylamide gel electrophoretic pattern of dsRNA species isolated from virus-infected tissue. (A) carnation ringspot virus, (B) red clover necrotic mosaic virus.

Ultrathin section of cytoplasmic crystal composed of virus particles from inoculated cowpea leaf. Bar represents 0.5 µm.

Section of infected Dianthus barbatus leaf showing nucleus containing: v, virus particles; t, tubules; and cr, protein crystal. Bar represents 500 nm.
References list for DPV: Carnation ringspot virus (308)
- Bercks & Querfurth, Phytopath. Z. 75: 215, 1972.
- Brown & Trudgill, Nematologica 30: 102, 1984.
- Bremer & Lahdenpera, Ann. Agric. Fenn. 20: 214, 1981.
- Dodds, Tremaine & Ronald, Virology 83: 322, 1977.
- Fritzsche & Schmelzer, Naturwissenschaften 54: 498, 1967.
- Fritzsche, Kegler, Thiele & Gruber, Arch. Phytopath. PflSchutz. 15: 177, 1979.
- Henriques & Morris, Virology 99: 66, 1979.
- Hiruki, Rao, Furuya & Figueiredo, J. gen. Virol. 65: 2273, 1984.
- Hollings & Stone, Ann. appl. Biol. 56: 73, 1965.
- Hollings & Stone, CMI/AAB Descr. Pl. Viruses 21, 3 pp., 1970.
- Hollings & Stone, CMI/AAB Descr. Pl. Viruses 181, 4 pp., 1977.
- Hollings, Stone & Bouttell, Ann. appl. Biol. 65: 299, 1970.
- Kalmakoff & Tremaine, Virology 33: 10, 1967.
- Kassanis, Ann. appl. Biol. 43: 103, 1955.
- Kegler & Kegler, Arch. Phytopath. PflSchutz. 17: 307, 1981.
- Kleinhempel, Gruber & Kegler, Acta phytopath. Acad. Sci. hung. 15: 107, 1980.
- Lane, in Nucleic Acids in Plants, vol. 2, p. 65, ed. T. C. Hall & J. W. Davies, Boca Raton, Florida, CRC Press: 237 pp., 1979.
- Lommel, Ph.D. Thesis, University of California, Berkeley, 1983.
- Lommel & Morris, Phytopathology 72: 955, 1982.
- Lommel, McCain & Morris, Phytopathology 72: 1018, 1982.
- Lommel, McCain, Mayhew & Morris, Pl. Dis. 67: 53, 1983.
- Lovisolo & Lisa, Fitopatol. Bras. 3: 219, 1978.
- Matthews, Intervirology 17: 1, 1982.
- Morris-Krsinich, Forster & Mossop, Virology 124: 349, 1983.
- Richter, Kleinhempel, Gruber & Kegler, Arch. Phytopath PflSchutz. 14: 411, 1978.
- Ronald & Tremaine, Phytopathology 66: 1302, 1976.
- Rudel, Querfurth & Paul, NachrBl. dt. PflSchutzdienst., Stuttg. 29: 59, 1977.
- Rybalko & Kharuta, Fiziol. i Biokhim. Kul't. Rast. (Physiol Biochem. cult. Pl.) 10: 535, 1978.
- Tremaine & Ronald, J. gen. Virol. 30: 299, 1976.
- Tremaine, Ronald & Valcic, Phytopathology 66: 34, 1976.
- Tremaine, Ronald & McGauley, Phytopathology 73: 1241, 1983.
- Tremaine, Ronald & McGauley, Phytopathology 74: 161, 1984.
- Trudgill, Brown & McNamara, Revue Nématol. 6: 133, 1983.
- Weintraub, Ragetli & Leung, Phytomorphology 25: 288, 1975.