Details of DPV and References
DPV NO: 249 July 1982
Family: Luteoviridae
Genus: Polerovirus
Species: Carrot red leaf virus | Acronym: CtRLV
Carrot red leaf virus
P. M. Waterhouse Scottish Crop Research Institute, Invergowrie, Dundee, Scotland
A. F. Murant Scottish Crop Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Stubbs (1948, 1952), Watson, Serjeant & Lennon (1964) and Waterhouse & Murant (1981a).
A virus with RNA-containing isometric particles c. 25 nm in diameter, transmitted by the aphid Cavariella aegopodii in the persistent (circulative) manner but not by inoculation with sap. It is apparently confined to phloem tissue, and infects only species in the family Umbelliferae. Distributed throughout the world, frequently in association with carrot mottle virus, which is dependent on it for transmission by aphids.
Main Diseases
In carrot (Daucus carota), the virus causes reddening and yellowing of leaves and some stunting of the plants. In nature, it commonly occurs in association with carrot mottle virus (Watson et al., 1964; Murant, 1974) to cause the disease known as ‘motley dwarf’, in which the reddening, yellowing and stunting is accentuated (see Fig. 1 of Murant, 1974) and there is a variable amount of mottling. The motley dwarf disease was first described and named by Stubbs (1948; 1952), but the existence of two virus components was not then recognized. The virus complex caused serious losses in spring-sown carrots and in carrot seed crops in Australia in the 1930s and 1940s (Stubbs, 1948). It has also caused serious yield losses in some years in Britain (Watson, 1960; Watson & Serjeant, 1964) and in the USA (Krass & Schlegel, 1974). A similar disease complex exists in parsley (Petroselinum crispum) in Britain (Frowd & Tomlinson, 1972) but the relationship of the causal viruses to carrot red leaf and carrot mottle viruses remains to be established (see below, under Strains).
Geographical Distribution
Carrot motley dwarf disease is reported from Australia (Stubbs, 1948; 1952), USA (Stubbs, 1956; Howell & Mink, 1974; Krass & Schlegel, 1974), Japan (Komuro & Yamashita, 1956), New Zealand (Anon., 1959), Europe (Watson, 1960; Heinze, 1968) and Canada (Murant, 1975). Both the carrot red leaf and carrot mottle virus components are presumably present in all these countries.
Host Range and Symptomatology
Occurs naturally in cultivated and wild carrot and is also reported from parsley (though see below under Strains), cow parsley (Anthriscus sylvestris), and hogweed (Heracleum sphondylium). Experimentally, it is restricted to a few species of Umbelliferae (Watson et al., 1964; P. M. Waterhouse & A. F. Murant, unpublished data), in which it typically induces stunting, reddening and yellowing. Transmitted by aphids but not by inoculation with sap. The following are useful test species.
- Diagnostic species
- Anthriscus cerefolium (chervil). Mild to severe stunting,
and curling of leaves, 10-16 days after inoculation (Fig. 1). Similar
symptoms occur when carrot mottle virus is also present, but the
stunting may be more severe, the leaflets being slightly more reduced
in size than with carrot red leaf virus alone (Fig. 1).
- Apium leptophyllum, syn. A. ammi (slender celery). Severe stunting and reddening, visible 7-14 days after inoculation. Similar symptoms occur when carrot mottle virus is also present.
- Coriandrum sativum (coriander). Severe stunting, bright yellowing and reddening of the whole plant, visible 12-16 days after inoculation (Fig. 2). Similar symptoms occur when carrot mottle virus is also present.
- Apium graveolens (celery) and Petroselinum crispum (parsley) are immune to most isolates (but see below under Strains); they can, however, be infected (symptomlessly) by carrot mottle virus.
- Apium leptophyllum, syn. A. ammi (slender celery). Severe stunting and reddening, visible 7-14 days after inoculation. Similar symptoms occur when carrot mottle virus is also present.
- Propagation species
- Daucus carota and Anthriscus cerefolium are
convenient for the maintenance of cultures; Anthriscus
cerefolium is used as a source of virus for purification.
- Assay species
- Anthriscus cerefolium, Apium leptophyllum and Coriandrum sativum are used in tests for transmission by aphids.
Strains
Few isolates have been compared serologically and most isolates described from carrot have similar host ranges. However, Frowd & Tomlinson (1972) described an isolate from parsley, a species which is immune to carrot isolates, and Ohki, Doi & Yora (1979) described an isolate from carrot in Japan which was unusual in infecting celery (Apium graveolens) and Cryptotaenia japonica. Although these isolates may well be host range variants of carrot red leaf virus, there is as yet no serological evidence that they are related to it.
Transmission by Vectors
Transmitted in the persistent (circulative) manner by the carrot-willow aphid, Cavariella aegopodii, but not by eight other species tested: Anuraphis tulipae, Acyrthosiphon pisum, Cavariella pastinacae, C. theobaldi, Hyadaphis foeniculi, Myzus persicae and two unidentified species of Macrosiphum (Stubbs, 1948; Watson et al., 1964; Murant, 1974; Waterhouse, 1981). The carrot virus from Japan (Ohki et al., 1979) was transmitted both by Cavariella aegopodii and by Semiaphis heraclei.
In tests with C. aegopodii, the minimum acquisition access period was about 30 min and the minimum inoculation access period about 2 min (Elnagar & Murant, 1978a); the minimum latent period was between 6 and 18 h (Watson et al., 1964; Elnagar & Murant, 1978a). Aphids retained the ability to transmit the virus after moulting (Stubbs, 1955; Watson et al., 1964; Elnagar & Murant, 1978a) and continued to transmit it after at least 12 serial daily transfers (Elnagar & Murant, 1978a). The virus was not transmitted to the progeny of viruliferous aphids and there is no evidence for multiplication of the virus in the vector (Elnagar & Murant, 1978a, 1978b). Aphids injected with extracts of viruliferous donor aphids transmitted carrot red leaf virus readily to test plants (Elnagar & Murant, 1978b).
Carrot red leaf virus acts as a helper for transmission of carrot mottle virus by aphids (Watson et al., 1964). In mixedly infected plants, some carrot mottle virus nucleic acid becomes packaged in coat protein of carrot red leaf virus (Waterhouse, 1981; Waterhouse & Murant, 1981b); these particles are transmitted by C. aegopodii in the same way as carrot red leaf virus particles and have the same vector relations (Elnagar & Murant, 1978a, 1978b).
Transmission through Seed
Not seed-transmitted in carrot (Stubbs, 1948; Krass & Schlegel, 1974; A. F. Murant, unpublished data) or in Anthriscus cerefolium (Waterhouse, 1981).
Serology
A rabbit injected intradermally with 100 µg purified virus yielded antiserum with a titre of 1/512 in double diffusion tests in agarose gel (Waterhouse & Murant, 1981a). A single line of precipitate is produced in such tests but they must be done with partially purified virus because the concentration of virus in plant extracts is too low. However, the virus can be detected in unconcentrated extracts from plants or groups of aphids by using enzyme-linked immunosorbent assay (ELISA) or immunosorbent electron microscopy (ISEM) (Waterhouse & Murant, 1981a). Analysis of antigen-antibody reactions by sucrose density gradient centrifugation is a useful technique for determining serological relationships (Waterhouse & Murant, 1981a).
Relationships
The properties of carrot red leaf virus (induction of reddening and yellowing symptoms, restriction to phloem tissue, non-transmissibility by mechanical inoculation, high vector specificity, persistent (circulative) relation with the aphid vector and possession of 25 nm isometric particles) place it in the luteovirus group. It is distantly serologically related to each of seven luteoviruses tested (Waterhouse & Murant, 1981a), most closely to barley yellow dwarf virus (RPV strain) and perhaps beet western yellows virus, more distantly to tobacco necrotic dwarf, potato leafroll and bean leafroll viruses, and very distantly to barley yellow dwarf virus (MAV strain) and soybean dwarf virus.
Stability in Sap
Infectivity of purified preparations survived after 1 month at -15°C (Waterhouse, 1981).
Purification
The following method is useful (Waterhouse & Murant, 1981a). Homogenise infected chervil plants in 0.1 M sodium citrate buffer, pH 6.0, containing 1% (w/v) thioglycerol (3 ml/g tissue). Centrifuge the resultant slurry at 3000 g for 15 min and discard the supernatant fluid. Resuspend the pellets, mainly fibrous material, in extraction buffer containing 0.02% (w/v) sodium azide and 1.5% (w/v) Driselase (a product that contains pectinase and cellulase enzymes); then shake the suspension at 28°C for 16-20 h. After incubation add Triton X-100 to 1% (v/v) and stir at room temperature for 30 min. Subject the suspension to two cycles of differential centrifugation: in each cycle, first centrifuge the preparation at 8000 g for 15 min and then float the supernatant fluid on a layer of 20% (w/v) sucrose occupying about one quarter of the tube, and centrifuge for 3.5 h at 140,000 g. Resuspend the pellets from high speed centrifugation in 0.006 M phosphate buffer, pH 7.0. The virus may be further purified by sucrose density gradient centrifugation. About 1 mg virus may be obtained from 1 kg leaf material.
Properties of Particles
The particles form a single sedimenting and buoyant density component (Fig. 4).
Sedimentation coefficient (s20,w): 104 S.
Buoyant density in CsCl: 1.403 g/cm3.
A260/A280: 1.62; Amax(260)/Amin(240): 1.22 (values not corrected for light-scattering).
Absorption coefficient (A260,0.1%,1 cm): c. 7 .0, estimated from the A260/A280 ratio and buoyant density.
Particle Structure
The particles are isometric, c. 25 nm in diameter, many appearing hexagonal in outline (Fig. 3).
Particle Composition
Nucleic acid: RNA, single-stranded, about 28% of particle weight, estimated from the buoyant density of the particles in CsCl. Electrophoresis in polyacrylamide gels in non-denaturing conditions revealed only one species, of M. Wt c. 1.8 x 106 (Waterhouse, 1981).
Protein: Protein of M. Wt c. 25,000 was observed
by electrophoresis in polyacrylamide/SDS gels. Three minor
slower-migrating components were found but their origin is unknown
(Waterhouse, 1981).
Relations with Cells and Tissues
The virus particles are confined to the phloem tissue; they occur especially in companion cells (Fig. 5) and sieve tubes, but also in phloem parenchyma cells (Murant & Roberts, 1979). Particles were found in the cytoplasm and vacuoles but not in the nuclei of infected cells. A similar tissue distribution was reported for the Japanese carrot virus studied by Ohki et al. (1979), who also observed cytoplasmic vesicular structures containing nucleic acid-like fibrils.
Notes
Carrot red leaf virus is the only definitive luteovirus known to infect species of Umbelliferae. The symptoms it causes in carrot, whether alone or in association with carrot mottle virus, are not easily confused with those caused by any other virus but may be mistaken for those of a deficiency disorder (Stubbs, 1948) or with those caused by carrot fly (Psila rosae). Insecticide treatments applied against carrot fly are also effective against Cavariella aegopodii. Carrot red leaf and carrot mottle viruses are therefore now uncommon in areas where routine treatments against carrot fly are employed. The viruses do not infect willow, the winter host of C. aegopodii, but survive between crops in weeds such as cow parsley or wild carrot, in volunteer carrots or in overwintering carrot seed crops (Watson & Serjeant, 1964; Howell & Mink, 1977a, 1977b). The virus may also be carried in roots transplanted into new areas for seed production (Stubbs, 1948; Howell & Mink, 1976). The epidemiology of motley dwarf disease was investigated by Watson & Serjeant (1964) and Howell & Mink (1977a, 1977b).
Figures

Chervil, Anthriscus cerefolium: (left) healthy, (upper right) infected with carrot red leaf virus, (lower right) infected with both carrot red leaf and carrot mottle viruses.

Coriander, Coriandrum sativum: (left) healthy, (right) infected with carrot red leaf virus.

Particles of carrot red leaf virus. Prepared by the method of Horne & Ronchetti (1974) with 2% ammonium molybdate, pH 6.5, and 2% uranyl acetate. Bar represents 100 nm. (From Waterhouse & Murant, 1981a).

Schlieren patterns: (above) moving boundary sedimentation in 0.006 M phosphate buffer, pH 7.0, at 20°C. Photograph taken 8 min after a speed of 30,000 rev./min was reached; (below) equilibrium banding after centrifugation for 16 h at 20°C at 44,000 rev./min in a CsCl solution of initial density 1.44 g/cm3. (From Waterhouse & Murant, 1981a).
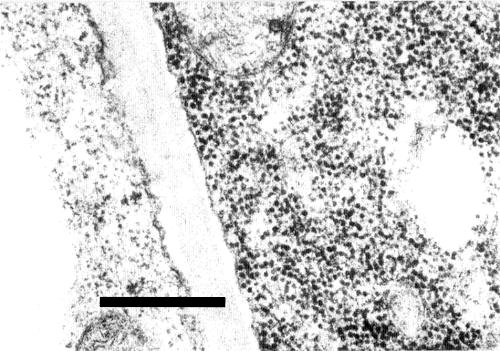
Particles of carrot red leaf virus in a companion cell from a thin section of an infected chervil leaf. The virus particles are readily distinguished from the ribosomes in the adjacent parenchyma cell which contains no virus-like particles. (From Murant & Roberts, 1979). Bar represents 500 nm.
References list for DPV: Carrot red leaf virus (249)
- Anon., Rep. Dep. scient. ind. Res. N.Z. 1959, 112 pp., 1959.
- Elnagar & Murant, Ann. appl. Biol. 89: 237, 1978a.
- Elnagar & Murant, Ann. appl. Biol. 89: 245, 1978b.
- Frowd & Tomlinson, Ann. appl. Biol. 72: 177, 1972.
- Heinze, Z. PflKrankh. Pflpath. PflSchutz 75: 513, 1968.
- Horne & Ronchetti, J. Ultrastruct. Res. 47: 361, 1974.
- Howell & Mink, Pl. Dis. Reptr. 58: 766, 1974.
- Howell & Mink, Pl. Dis. Reptr 60: 1047, 1976.
- Howell & Mink, Pl. Dis. Reptr 61: 217, 1977a.
- Howell & Mink, Pl. Dis. Reptr 61: 841, 1977b.
- Komuro & Yamashita, Ann. phytopath. Soc. Japan 20: 155, 1956.
- Krass & Schlegel, Phytopathology 64: 151, 1974.
- Murant, CMI/AAB Descr. Pl. Viruses 137, 4 pp., 1974.
- Murant, Can. Pl. Dis. Surv. 55: 103, 1975.
- Murant & Roberts, Ann. appl. Biol. 92: 343, 1979.
- Ohki, Doi & Yora, Ann. phytopath. Soc. Japan 45: 74, 1979.
- Stubbs, Aust. J. sci. Res. B 1: 303, 1948.
- Stubbs, Aust. J. sci. Res. B 5: 399, 1952.
- Stubbs, J. Aust. Inst. agric. Sci 21: 267, 1955.
- Stubbs, Pl. Dis. Reptr 40: 763, 1956.
- Waterhouse, Ph.D. Thesis, University of Dundee, 244 pp., 1981.
- Waterhouse & Murant, Ann. appl. Biol. 97: 191, 1981a.
- Waterhouse & Murant, Abstr. 5th int. Congr. Virol., Strasbourg, France, 1981: 212, 1981b.
- Watson, Pl. Path. 9: 133, 1960.
- Watson & Serjeant, Ann. appl. Biol. 53: 77, 1964.
- Watson, Serjeant & Lennon, Ann. appl. Biol. 54: 153, 1964.