Details of DPV and References
DPV NO: 339 September 1988
Family:
Genus:
Species: | Acronym:
Includes viruses in the genera Luteovirus and Polerovirus and those in the family Luteoviridae not yet assigned to a genus.
Luteovirus group
P. M. Waterhouse Division of Plant Industry, CSIRO, Canberra, Australia
F. E. Gildow Department of Plant Pathology, Pennsylvania State University, USA
G. R. Johnstone Tasmania Department of Agriculture, Hobart, Australia
Contents
- Type Member
- Main Characteristics
- Members
- Geographical Distribution
- Transmission by Vectors
- Ecology and Control
- Relations with Cells and Tissues
- Properties of Particles
- Genome Properties
- Replication
- Satellite
- Defective-Interfering RNA
- Relationships
- Notes on Tentative Members
- Affinities with Other Groups
- Notes
- Acknowledgements
- Figures
- References
Type Member
Barley yellow dwarf virus - strain MAV.
Main Characteristics
Isometric particles c. 25 nm in diameter sedimenting as a single component at c. 104-118 S, and with a buoyant density in CsCl of c. 1.40 g/cm3. The protein shell is composed of one polypeptide species of M. Wt c. 24 x 103 and the genome is a single molecule of positive sense, single-stranded RNA (M. Wt c. 2.0 x 106). Luteovirus particles have high A260/A280 ratios of c. 1.6-1.9, are moderately stable (thermal inactivation points of 45-75°C), and are strongly immunogenic. Most members are serologically inter-related.
The name luteovirus (Latin, luteus = yellow) was coined
because infected plants tend to show yellowing
symptoms. Other characteristic symptoms are reddening, leaf curling,
leaf rolling and brittleness. The concentration of virus particles
within the plant is low (less than 100 µg/l of sap) and
particles have been observed only in phloem tissues. Luteoviruses
are transmitted neither by mechanical inoculation nor through seed,
but are transmitted by aphids in a persistent (circulative,
non-propagative) manner. Most members show a high degree of vector
specificity and some luteoviruses can act as helpers for the
aphid-transmission of associated viruses. The International
Committee on Taxonomy of Viruses defined the luteovirus group in
1975 (Shepherd et al., 1976). It has since been reviewed by
Duffus (1977b),
Francki et al. (1985) and
Rochow &
Duffus (1981).
Members
Definitive and tentative members of the group are listed in Table 1. Those listed as possible or probable members resemble definitive luteoviruses in symptom expression and in their aphid transmission characteristics but are not sufficiently well studied to be classified unequivocally. Until recently, subterranean clover stunt virus was tentatively classified as a luteovirus. However, we exclude it from the group because its genome appears to be single-stranded DNA (Chu & Helms, 1987).
Table 1(a) Properties of definitive members of the luteovirus group (probable strains or synonyms are shown in italics and preceded by = )
| Main host family† |
Main vector | Particle diam. (nm) |
Genome RNA M. Wt (× 10-6) |
Particle protein M. Wt (× 10-3) |
A260/A280 | s20,w | Density in CsCl (g/cm3) |
DPV no. or reference |
|
| Barley yellow dwarf (BYDV)-MAV | |||||||||
| Gr | Sitobion avenae | 26, 30, 24T | 1.85 | 23.5 | 1.88-1.92 | 115-118 | 1.39 | 32; a-d | |
| = (BYDV-PAV) | |||||||||
| Gr | S. avenae, Rhopalosiphum padi | 26, 24T | 2.0 | 24.4 | 1.76 | 115 | 1.40 | 32; a-d | |
| = (BYDV-SGV) | |||||||||
| Gr | Schizaphis graminum | 24T | 32 | ||||||
| Barley yellow dwarf (BYDV)-RPV | |||||||||
| Gr | R. padi | 26, 30, 24T | 1.85-2.0 | 24.4 | 1.71-1.79 | 115-118 | 32; a-d | ||
| = Rice guillaume (RGV) | |||||||||
| Gr | R. padi | 28 | e | ||||||
| = BYDV-RMV | |||||||||
| Gr | R. maidis | 24T | 32 | ||||||
| Bean leaf roll (BLRV) | |||||||||
| Le | Acyrthosiphon pisum | 27, 23T | 2.4 | 23 | 1.83 | 1.32* | 286 | ||
| = Legume yellows (LYV) | |||||||||
| Le | Acyrth. pisum | f | |||||||
| = Michigan (lucerne) alfalfa (MiAV) | |||||||||
| Le | Acyrth. pisum | g | |||||||
| = Pea leaf roll | |||||||||
| Le | Acyrth. pisum | h | |||||||
| Beet western yellows (BWYV) | |||||||||
| Am, Ch, Co, Cr, Le, So | Myzus persicae | 27, 26T | 1.9 | 24 | 89; i; j | ||||
| = Beet mild yellowing (BMYV) | |||||||||
| Ch, Cr | M. persicae | 26 | 2.0 | 24-25.4 | 1.8 | 116 | 1.42 | k, l | |
| = Malva yellows (MYV) | |||||||||
| Ch, Co, Cr, Ma, So | M. persicae | m | |||||||
| = Turnip mild yellows (TuYV) | |||||||||
| Co, Cr | M. persicae | n | |||||||
| Carrot red leaf (CRLV) | |||||||||
| Um | Cavariella aegopodii | 25 | 1.8 | 25 | 1.62 | 104 | 1.39 | 249 | |
| Groundnut rosette assistor (GRAV) | |||||||||
| Le | Aphis craccivora | 28-30 | 2.1 | 24 | 1.86 | 115 | 1.34* | o, p | |
| Indonesian soybean dwarf (ISDV) | |||||||||
| Le | Aphis glycines | 26, 24T | q | ||||||
| Potato leafroll (PLRV) | |||||||||
| Am, So | M. persicae | 24-25 | 2.0 | 26.3 | 1.78 | 115-127 | 1.39; 1.32* | 36; 291 | |
| = Solanum yellows (SYV) | |||||||||
| So | M. persicae | 28-30 | r | ||||||
| = Tomato yellow top (TYTV) | |||||||||
| So | M. persicae | 24 | 25.7 | 1.34* | s | ||||
| Soybean dwarf (SDV) | |||||||||
| Le | Aulacorthum solani | 25 | 22 | 1.96-1.90 | 179 | ||||
| = Subterranean clover red leaf (SCRLV) | |||||||||
| Le | Aul. solani | 25 | 2.0 | 22.6 | 1.85 | 114 | t, u | ||
| = Strawberry mild yellow edge (SMYEV) | |||||||||
| Fragaria spp. | Chaetosiphon fragaefolii | 23 | v, w | ||||||
| Tobacco necrotic dwarf (TNDV) | |||||||||
| So | M. persicae | 25 | 2.0 | 25.7 | 1.8 | 115 | 234 | ||
Table 1(b) Probable and possible members of the luteovirus group
| Probable members | Main host family† | Main vector species | Comments | Reference |
| Banana bunchy top (BBTV) | Musa spp. | Pentolonia nigranervosa | Double-stranded RNA profile typical of a luteovirus | x |
| Milk vetch dwarf | Le | A. craccivora | Isometric particles purified and seen in thin sections of phloem tissue | y |
| Millet red leaf | Gr | R. maidis, S. avenae, Sch. graminum | Possibly a form of BYDV | z |
| Physalis mild chlorosis | Cr, So | M. persicae | Possibly a form of BWYV | aa |
| Tobacco vein distorting (TVDV) | So | M. persicae | Acts as a helper virus | bb |
| Tobacco yellow vein assistor (TYVAV) | So | M. persicae | Acts as a helper virus | cc |
| Tobacco yellow net | So | M. persicae | dd | |
| Possible members | ||||
| Beet yellow net | Ch | M. persicae | ee | |
| Celery yellow spot | Um | R. conii | ff | |
| Cotton anthocyanosis | Ma | Aphis gossypii | gg | |
| Filaree red leaf | Geraniaceae | Acyrth. pe!argonii | hh | |
| Grapevine ajinashika | Vitis spp. | Vector unknown | ii | |
| Physalis vein blotch | So | M. persicae | jj | |
| Raspberry leaf curl | Rosaceae | Aphis rubicola | kk |
T= Measurement of particles seen in thin sections. * = Buoyant density in caesium sulphate solutions. † Abbreviations for family names: Am Amaranthaceae; Ch Chenopodiaceae; Co Compositae; Cr = Cruciferae; Gr = Gramineae; Le = Leguminosae; So = Solanaceae; Ma = Malvaceae; Um = Umbelliferae.
References: a: Brakke & Rochow, 1974;
b: Scalla & Rochow, 1977;
c: Paliwal, 1978;
d: Hammond et al., 1983;
e: Amici et al., 1978;
f: Duffus, 1979;
g: Thottappilly et al., 1977;
h: Hampton, 1983;
i: Falk et al., 1977;
j: Falk & Duffus, 1984;
k: Chevallier et al., 1983;
l: Govier, 1985;
m: Costa et al., 1959;
n: Watson, 1964;
o: Casper et al., 1983;
p: Rajeshwari & Murant, 1988;
q: Iwaki et al., 1980;
r: Milbrath & Duffus, 1978;
s: Thomas, 1984;
t: Johnstone et al., 1982;
u: Ashby & Kyriakou, 1983;
v: Martin & Converse, 1985;
w: Spiegel et al., 1986;
x: Dale et al., 1986;
y: Inouye et al., 1979;
z: Yu et al., 1957;
aa: MacKinnon, 1965;
bb: Smith, 1946;
cc: Adams & Hull, 1972;
dd: Sylvester, 1954;
ee: Sylvester, 1948;
ff: Freitag & Severin, 1945;
gg: Costa, 1957;
hh: Frazier, 1951;
ii: Namba et al., 1979;
jj: MacKinnon & Lawson, 1966;
kk: Bennett, 1930.
Geographical Distribution
Luteoviruses have been described from most parts of the world including tropical, sub-tropical and temperate regions. Many luteoviruses, including BYDV, BWYV, CRLV and PLRV, are widespread but others, such as TNDV, have restricted distributions. With the exception of BWYV and close serological relatives, all the definitive members have natural host ranges largely restricted to one plant family (Table 1).
Transmission by Vectors
Luteoviruses are transmitted in a persistent (circulative, non-propagative) manner. Aphids acquire and transmit luteoviruses while feeding on phloem sieve tube elements of host plants. Minimum feeding access times reported for aphids to acquire or inoculate different luteoviruses range from 0.1-4.0 h and 0.2-1.0 h respectively (Cockbain & Costa, 1973; Duffus, 1960, 1979; Elnagar & Murant, 1978; Thottappilly et al., 1977). These reported times include the time required for the aphids’ stylets to penetrate to the phloem tissues. Mean times for stylet penetration of sieve tube elements in oats were 200, 60 and 32 min for Schizaphis graminum, Rhopalosiphum padi and Sitobion avenae respectively (Montllor & Gildow, 1986; Scheller & Shukle, 1986). Electronic monitoring of Myzus persicae on potato showed that aphids whose stylets had penetrated and remained in sieve elements for a minium of 1-2 min subsequently became able to transmit PLRV (Leonard & Holbrook, 1978). Efficient transmission of most luteoviruses requires acquisition and inoculation feeding access times each of 24 h. The minimum latent period (usually defined as the time from the start of the acquisition feeding period until the insect becomes able to infect plants with the virus) is normally between 12 and 24 h (Duffus, 1977b; Elnagar & Murant, 1978; Rochow, 1963). Median latent periods (LP5O, time for 50% of the aphids to begin to transmit) are 35-50 h at 15-20°C for BYDV-RPV, 44-65 h at 15-20°C for BYDV-MAV (Van der Broek & Gill, 1980) and 30-49 h at 20-25°C for PLRV (Eskandari et al., 1979). Factors influencing luteovirus transmission were described by Sylvester (1980).
Aphids that have acquired a luteovirus do not cease to transmit
it after moulting and studies of BYDV transmission by R. padi
(Gildow, 1985) indicate that luteovirus particles pass by cellular
transport through the hindgut into the aphid’s haemocoel. The
particles then circulate in the haemolymph throughout the aphid.
Current evidence indicates that luteoviruses do not replicate in
their vectors (although Weidemann (1982) suggested that they may do
so to a small extent), but aphids that have begun to transmit a
luteovirus may continue to do so for 2-3 weeks without
re-acquisition. Ultrastructural studies on BWYV, BYDV and PLRV
suggest that they are transmitted following endocytosis of virus
particles from the haemocoel into the aphid’s accessory salivary
gland. The particles are then transported within coated vesicles
through the salivary gland secretory cell, and released by
exocytosis into the salivary duct (Gildow & Rochow, 1980a;
Gildow 1982, 1987).
A characteristic feature of the luteoviruses is their high degree
of vector specificity (Table 1).
Each luteovirus is transmitted
efficiently by only one or a few species of aphid. Vector
specificity is believed to result from interactions between the
particle protein and membrane surfaces in the aphid’s accessory
salivary gland; these interactions regulate transport of virus
particles to the salivary duct (Gildow & Rochow, 1980b).
Vector specificity is not absolute, however, and some luteoviruses
are transmitted inefficiently by species other than the main vector
(Rochow, 1969). Transmission efficiency has also been reported to
vary with the age of the aphid (Gill, 1970; Upreti & Nagaich,
1971; Zhou & Rochow, 1984), and among distinct populations or
clones of a single aphid species (Bjorling & Ossiannilsson,
1958; Cockbain & Costa, 1973; Gildow & Rochow, 1983; Rochow,
1960). Genetic variability of aphids, as well as the viruses,
evidently influences vector specificity and transmission efficiency.
Ecology and Control
Some luteoviruses may be disseminated with infected planting material, for example BBTV with banana suckers, PLRV with potato tubers, and SMYEV with strawberry runners. Clean stock schemes are operated in many parts of the world to control these viruses (Allen, 1983; Harrison, 1984; Mullin et al., 1974). Most other luteoviruses cause diseases in annual crops and are introduced each year by their aphid vectors. The viruses spread into the annual crops from reservoirs which include volunteer plants from the previous year's crop, other crops, alternative perennial hosts and weeds. In some instances, infective vectors may be transported long distances in wind currents (Fargette et al., 1982). Weeds are a particularly important source of BWYV infection, and perennial grasses and legumes are important reservoirs of BYDV and legume-infecting luteoviruses (SDV, BLRV, BWYV) respectively. The relative importance of secondary spread by apterae within a crop and of primary introductions by alatae varies with the virus, the vector species, the crop species and the climatic conditions. Organophosphate aphicides help control secondary spread of luteoviruses but are ineffective against primary introductions. In contrast, synthetic pyrethroids have a rapid knock-down property that can be useful in decreasing primary spread (Gibson et al., 1982; Johnstone, 1984). Other control approaches have included judicious choice of sowing date in relation to aphid flight activity (Stubbs, 1948), elimination of virus sources (Duffus, 1971), adoption of crop-free periods (Duffus, 1977a), crop production in isolation (Shepard & Claflin, 1975), breeding for various forms of virus resistance or tolerance (Barker & Harrison, 1985; Jedlinski et al., 1977; Schalter et al., 1963), breeding for resistance to aphid feeding (Gibson, 1977) and biological control of the aphid vector (Stubbs et al., 1983).
A number of luteoviruses occur in the field as part of a
complex, either with other luteoviruses, such as BWYV and SDV in legumes (Johnstone & Duffus, 1984) and various BYDV serotypes
in cereals (Rochow & Muller, 1974), or with members of other
virus groups, as with the virus complexes found in strawberries
(Prentice, 1946). Some viruses depend on luteoviruses for their
aphid transmission (Table 2), for example carrot mottle virus
depends on CRLV, and groundnut rosette virus depends on GRAV.
These pairs of viruses occur as complexes that seem to be very
stable and may show a synergistic effect with regard to symptoms.
Table 2. Luteoviruses which have a helper activity and their associated dependent virus
| Helper virus | Vector | Dependent virus | Reference |
| BYDV-RMV | Rhopalosiphum maidis | BYDV-MAV | Rochow, 1975 |
| BYDV-RMV | R. maidis | BYDV-RPV | Rochow, 1982 |
| BYDV-RPV | R. padi | BYDV-MAV | Rochow, 1970 |
| BYDV-RPV | R. padi | BYDV-RMV | Rochow, 1982 |
| BYDV-RPV | R. padi | BYDV-SGV | Rochow, 1982 |
| BYDV-PAV | R. padi | BYDV-RMV | Rochow, 1982 |
| BYDV-PAV | R. padi | BYDV-MAV | Rochow, 1982 |
| BLRV | Acyrthosiphon pisum | Bean yellow vein-banding | Cockbain, 1978; Cockbain et al., 1986 |
| BWYV | Myzus persicae | Lettuce speckles mottle | Falk et al., 1979 |
| BWYV* | M. persicae | Carrot mottle | Waterhouse & Murant, 1983 |
| CRLV | Cavariella aegopodii | Carrot mottle | Watson et al., 1964 |
| GRAV | Aphis craccivora | Groundnut rosette | Hull & Adams, 1968 |
| GRAV* | A. craccivora | Tobacco yellow vein | Adams & Hull, 1972 |
| PLRV* | M. persicae | Carrot mottle | Waterhouse & Murant, 1983 |
| TVDV* | M. persicae | Tobacco yellow vein | Adams & Hull, 1972 |
| TVDV | M. persicae | Tobacco mottle | Smith, 1945; 1946 |
| TYVAV | M. persicae | Tobacco yellow vein | Adams & Hull, 1972 |
| TuYV | M. persicae | Lettuce speckles mottle | Falk et al., 1979 |
* = Shown to act as a helper experimentally but not known to do so in nature.
Relations with Cells and Tissues
Luteoviruses are tissue-specific and replicate in phloem cells of plant hosts. Virus particles are detected most readily in sieve elements, companion cells, and occasionally in phloem parenchyma. Particles were found only in phloem transfer cells of SCRLV-infected subterranean clover (Jayasena et al., 1981). With one exception (Gill & Chong, 1981), luteoviruses have not been observed in epidermal, mesophyll or xylem tissues of infected plants. Evidence suggests that luteoviruses spread rapidly from cell to cell longitudinally through the vascular bundle; however, lateral spread is slow and not all phloem cells of a bundle may be infected (Jensen, 1969; Murant & Roberts, 1979). The primary symptom of infection is necrosis of the phloem (Esau, 1957), which spreads from the initially inoculated sieve element to adjacent cells. Phloem necrosis is associated with resistance to translocation of plant metabolites, loss of chlorophyll, increased respiration, and decreased growth (Goodman et al., 1965; Jensen, 1972; Orlob & Arny, 1961).
Ultrastructural examinations of infected plants have been reported
for several luteoviruses (D’Arcy & De Zoeten, 1979; Esau &
Hoefert, 1972; Faoro et al., 1978; Jayasena et al.,
1981; Jensen, 1969; Kojima et al., 1969; Kubo, 1981; Murant
& Roberts, 1979; Shepardson et al., 1980). The most
detailed study was that of Gill & Chong (1979) on BYDV isolates
infecting oats, and showed that at least two distinct sequences of
events may be involved in replication of viruses assigned to the
luteovirus group. Based on cytopathological ultrastructure involving
alterations of the nucleus, site of virus accumulation and types of
vesicles produced, these authors suggested dividing BYDV isolates
into subgroup 1 (MAV, PAV, SGV) and subgroup 2 (RPV, RMV). In oats
infected with isolates of subgroup 1, virus particles seem to move
from sites of initial inoculation in sieve elements through
plasmodesmata into adjacent companion cells. Densely staining
filaments (2-4 nm diameter) and single membrane-bound vesicles
(50-230 nm diameter) form in the cytoplasm near the plasmodesmata.
Filaments are later observed within the nucleus, which becomes
distorted and begins to deteriorate soon after infection. Virus
particles are first observed to occur in the cytoplasm, suggesting
a cytoplasmic site for assembly. At about this time the
mitochondria, plastids and ribosomes begin to disintegrate.
Isolates in subgroup 2 initiate infection of companion cells in a
similar manner; however, the vesicles produced are bounded by a
second membrane which is continuous with the endoplasmic reticulum.
A second type of membrane system (composed of tubules) proliferates,
and the dense filaments are rare in the cytoplasm and do not occur
in the nucleus. The nucleus does not deteriorate and virus particles
are first observed surrounding the nucleolus within the nucleus. In
addition, extensive wall thickenings develop in the infected
parenchyma cells.
Ultrastructural changes induced by BWYV and PLRV seem to be
similar to those caused by BYDV isolates of subgroup 2 (Esau &
Hoefert, 1972; Shepardson et al., 1980). However,
cytopathological effects vary with host species and virus isolate
(D'Arcy & De Zoeten, 1979; Gill & Chong, 1981; Kojima &
Yanase, 1984).
Properties of Particles
Particles: Properties of luteovirus particles are summarised in Table 1. When negatively stained, most luteovirus particles have a diameter of c. 25-28 nm; they are usually angular in outline and not penetrated by stain. Thermal inactivation points range from 45°C to 75°C. The particle mass of BMYV is 6.5 x 106, as determined by optical diffraction and scanning transmission electron microscopy (Chevallier et al., 1983). The protein subunits of a luteovirus particle are probably in a T = 3 arrangement: the M. Wt of 180 protein subunits plus the M. Wt of the RNA gives a total very close to that obtained for the whole particle mass of BMYV. Hewings & D’Arcy (1983) reported that purified preparations of a legume isolate of BWYV contained not only the usual nucleoprotein particles but also particles made of the viral protein subunits but devoid of nucleic acid. The isoelectric point of TNDV particles is 5.3 (Kubo, 1981).
Nucleic acid: The genomes of all luteoviruses so far analysed consist of single-stranded RNA with a M. Wt of c. 2.0 x 106. The genome of PLRV was at first reported to be double-stranded DNA (Sarkar, 1976) but this has not been confirmed in more recent work (Rowhani & Stace-Smith, 1979; Takanami & Kubo, 1979b). The RNA molecules of PLRV, BYDV and CRLV each represent 28-30% of the particle weight (Harrison, 1984; Paliwal, 1978; Waterhouse & Murant, 1982). One isolate of BWYV, but not others, has a second RNA component (M. Wt 0.86 x 106) which was found to be associated with increased virulence (Falk & Duffus, 1984). However, this RNA component is probably not subgenomic (Falk & Anderson, 1987). The RNA extracted from TNDV particles also included two small RNA molecules (M. Wt 1 x 105), but their origin and significance is presently unknown (Takanami & Kubo, 1979a,1979b).
An isolate of BYDV-RPV studied by Miller et al.
(1987) has an associated small circular self-cleaving
single-stranded RNA (351 nt) which has no homology with the viral
genome and therefore appears to be satellite RNA.
Coat protein: The coat protein subunits of luteoviruses so far studied are all of M. Wt c. 24,000. The coat protein subunit of BLRV was originally reported to be c. 32,500 (Ashby & Huttinga, 1979) but a more recent estimate is c. 23,000 (R. O. Hampton, cited by Ashby, 1984). The amino acid composition of the coat protein subunits has been determined for only three luteoviruses: TNDV (Kubo, 1981), BYDV-PAV (P. M. Waterhouse & A. A. Kortt, unpublished data) and SDV (P. M. Waterhouse & A. A. Kortt, unpublished data):
lys
his
arg
asp
thr
ser
glu
pro
gly
ala
cys
val
met
ile
leu
try
phe
trp
TNDV:
10
4
21
20
12
24
16
9
23
12
3
13
2
10
23
6
9
2
SDV:
15
4
17
23
15
23
19
10
24
16
1
12
1
13
13
6
11
0
BYDV:
12
2
19
19
17
19
15
8
23
17
1
12
3
11
9
5
4
nd
The relatively low amounts of aromatic amino acids (mainly tyrosine and tryptophan) in the coat protein subunits of TNDV, SDV and BYDV may well account for the unusually high A260/A280 ratios observed for particle preparations of these and some other luteoviruses.
Genome Properties
The RNA of PLRV (Mayo et al., 1982) is covalently bound to a genome-linked protein (M. Wt 7000) and has no substantial polyadenylate sequence. The luteovirus genome is probably positive sense: viral polypeptides have been synthesised by in vitro translation of the encapsidated RNA of PLRV and SDV. Translation of PLRV RNA in rabbit reticulocyte lysates gave polypeptides of M. Wt c. 125,000 and 71,000 (Mayo et al., 1982). In wheat germ extracts these two and an additional polypeptide (M. Wt 29,000) were obtained (Mayo & Barker, 1984). SDV RNA gave one predominant translation product (M. Wt c. 45,000-50,000) with either the rabbit reticulocyte or the wheat germ systems (Anon., 1984). None of the polypeptides produced from translation of either RNA was precipitated by antiserum against the homologous coat protein.
The complete nucleotide sequence of a BYDV-PAV serotype has been
determined and its genome organization inferred from open reading
frames (Miller et al., 1988a,1988b). The gene nearest to
the 5' end encodes the RNA-dependent RNA polymerase, the coat protein
gene is located near the middle of the genome and there is a
possibility of translational readthrough from the coat protein
gene to yield a 69 kd protein.
The nucleotide sequence of a satellite RNA associated with an
isolate of BYDV-RPV is also known (Miller et al., 1987). It
has been inferred from the sequence that the satellite RNA has a
high degree of secondary structure and is not translated.
Replication
Little is known about the replication of luteoviruses. Although luteovirus particles are found only in phloem tissues, PLRV, TNDV and BYDV can replicate in mesophyll protoplasts (Barnett et al., 1981; Takanami & Kubo, 1979a) and SDV can replicate in soybean protoplasts derived from suspension cultures (Waterhouse & Gerlach, 1984). Furthermore, TNDV can replicate in inoculated epidermal cells (Imaizumi & Kubo, 1980). Therefore, the restriction of luteoviruses to cells of the phloem tissues is not because other cells are incapable of supporting their replication.
Subgenomic RNA species have been detected in nucleic acid
extracts from PLRV-infected potato (M. Wt 1 x 106;
Barker et al., 1984) and from BYDV-PAV -infected oats
(2.8 and 0.8 kb; Gerlach et al., 1987). Five species of
double-stranded RNA have been extracted from tissue infected with
BYDV-RPV (Gildow et al., 1983) or BWYV (Falk & Duffus,
1984) and four species from tissue infected with BYDV-MAV (Gildow
et al., 1983). This information and the incomplete
translation of PLRV or SDV RNA in vitro suggests that
luteoviruses may be translated via subgenomic messenger RNA species.
Relationships
The definitive luteoviruses form a network of serological relationships (Fig. 1). BWYV is directly serologically related to almost all definitive members. Luteoviruses seem to form a serological continuum but with a degree of clustering. Thus BWYV, BMYV, MYV and TuYV form a cluster which we have grouped as isolates of BWYV; similarly BLRV, LYV and MiAV form a cluster, as do PLRV, SYV and TYTV plus possibly TNDV. Also, SDV and SCRLV are very close. These same clusters are also seen when examining host range and vector specificity (Table 1). Paradoxically, BYDV isolates form not one cluster but two, one containing MAV, PAV and SGV, and the other containing RPV and RMV. This division is also supported by cytopathology (Gill & Chong, 1979), dsRNA profile (Gildow et al., 1983), nucleic acid hybridization (Waterhouse et al., 1986) and cross-protection studies (Jedlinski & Brown, 1965) and we suggest that these two subgroups should be regarded as two different viruses, BYDV-MAV and BYDV-RPV. Little work has been done on the inter-relationship of luteoviruses at the nucleic acid level. However, cross hybridization has been detected between regions of the BYDV-PAV genome and regions of the BYDV-RPV, SDV, PLRV and BWYV genomes (Waterhouse et al., 1986).
Affinities with Other Groups
Apart from the luteoviruses, the only other plant virus that is transmitted in a circulative, non-propagative, manner is pea enation mosaic virus (considered to form a monotypic group). There are many similarities between PEMV and luteoviruses in vector relations, particle morphology and composition, and in the ability to act as a helper virus. However, PEMV has at least two RNA components and is manually transmissible.
Sequence homology has been detected in the polymerase gene of
BYDV and that of carnation mottle virus and in the coat protein gene
of BYDV and that of southern bean mosaic virus (Miller
et al., 1988a, 1988b).
A number of luteoviruses can assist the aphid-transmission of
dependent viruses such as carrot mottle virus (Table 2). It has been shown for two
of these dependent viruses that their genomic RNA species can become
encapsidated in the coat protein of the helper virus
(Falk et al., 1979; Waterhouse & Murant, 1983), and this
is thought to explain how they become transmissible by the aphid
vector of the helper virus. It has also been shown that the
dependent virus can change its helper virus and consequently its
aphid vector (Adams & Hull, 1972; Waterhouse & Murant, 1983).
Figures
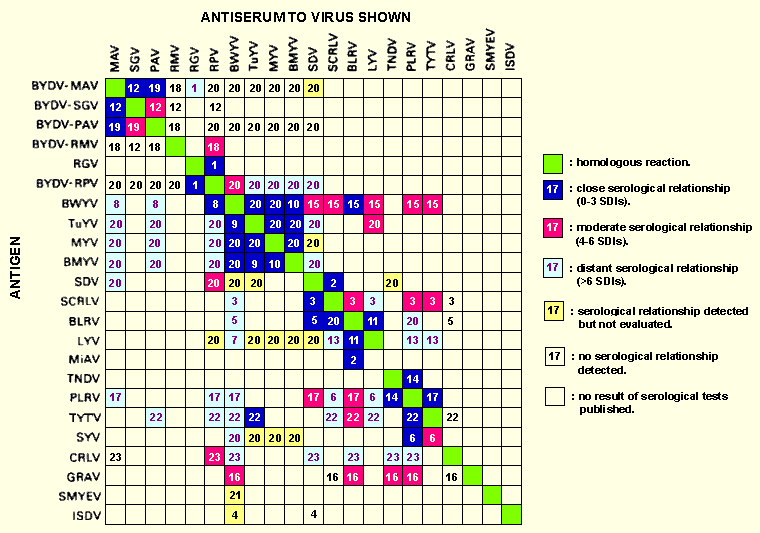
Serological relationships among luteoviruses.
Where possible SDIs have been used to determine the degree of
relationship but in some cases qualitative assessments have been
used. Numbers refer to the references listed below. The
strong serological relationship between TuYV antiserum and PLRV
antigen (22) may be due to the antiserum containing some antibodies
against PLRV.
References:
(1) Amici et al., 1978;
(2) Ashby, 1984;
(3) Ashby & Kyriakou, 1983;
(4) Ashby & Johnstone, 1985;
(5) Ashby & Huttinga, 1979;
(6) J. E. Duffus, unpublished data;
(7) Duffus, 1979;
(8) Duffus & Rochow, 1978;
(9) Duffus & Russell, 1972;
(10) Duffus & Russell, 1975;
(11) Duffus & Thottapilly, 1983;
(12) Hu et al., 1985;
(13) Johnstone, 1983;
(14) Kubo & Takanami, 1978;
(15) Kyriakou et al., 1983;
(16) Casper et al., 1983;
Reddy et al., 1985;
Rajeshwari & Murant, 1988;
(17) Roberts et al., 1980;
(18) Rochow & Carmichael, 1979;
(19) Rochow & Duffus, 1978;
(20) Rochow & Duffus, 1981;
(21) Spiegel et al., 1986;
(22) Thomas, 1984;
(23) Waterhouse & Murant, 1981.
References list for DPV: Luteovirus group (339)
- Adams & Hull, Ann. appl. Biol. 71: 135, 1972.
- Allen, in Plant Virus Epidemiology, p. 51, eds R. T. Plumb & J. M. Thresh, Blackwell, Oxford, 377 pp., 1983.
- Amici, Faoro, Osler & Tornaghi, Riv. Patol. veg., Pavia, Ser. IV 14: 127, 1978.
- Anon., Rep. Commonw. scient. ind. Res. Org. Div. Pl. Ind., 1983: 82, 1984.
- Ashby, CMI/AAB Descr. Pl. Viruses 286, 5 pp., 1984.
- Ashby & Huttinga, Neth. J. Pl. Path. 85: 113, 1979.
- Ashby & Johnstone, Australas. Pl. Path. 14: 2, 1985.
- Ashby & Kyriakou, N. Z. Jl agric. Res. 25: 607, 1983.
- Barker & Harrison, Ann. appl. Biol. 107: 205, 1985.
- Barker, Mayo & Robinson, Rep. Scott. Crop Res. Inst., 1983: 194, 1984.
- Barnett, Hammond & Lister, J. gen. Virol. 57: 397, 1981.
- Bennett, Phytopathology 20: 787, 1930.
- Bjorling & Ossiannilsson, Socker 14: 1, 1958.
- Brakke & Rochow, Virology 61: 240, 1974.
- Casper, Lesemann, Reddy, Rajeshwari, Misari & Subbarayudu, Phytopath. Z. 108: 12, 1983.
- Chevallier, Engel, Wurtz & Putz, J. gen. Virol. 64: 2289, 1983.
- Chu & Helms, Abstr. 7th int. Congr. Virol., Edmonton, Canada, 1987: 233 (abstr. no. R36.1), 1987.
- Cockbain, Rep. Rothamsted exp. Stn, 1977: 221, 1978.
- Cockbain & Costa, Ann. appl. Biol. 73: 167, 1973.
- Cockbain, Jones & Woods, Ann. appl. Biol. 108: 59, 1986.
- Costa, Phytopath. Z. 28: 167, 1957.
- Costa, Duffus & Bardin, J. Am. Soc. Sugar Beet Technol. 10: 371, 1959.
- D’Arcy & De Zoeten, Phytopathology 69: 1194, 1979.
- Dale, Phillips & Parry, J. gen. Virol. 67: 371, 1986.
- Duffus, Phytopathology 50: 389, 1960.
- Duffus, A. Rev. Phytopath. 9: 319, 1971.
- Duffus, Calif Agric. 31(10): 18, 1977a.
- Duffus, in Aphids as Virus Vectors, p. 361, eds K. Harris & K. Maramorosch, Academic Press, New York, 559 pp., 1977b.
- Duffus, Phytopathology 69: 217, 1979.
- Duffus & Rochow, Phytopathology 68: 45, 1978.
- Duffus & Russell, Phytopathology 62: 1274, 1972.
- Duffus & Russell, Phytopathology 65: 811, 1975.
- Duffus & Thottapilly, in Hampton, Pl. Dis. 67: 1306, 1983.
- Elnagar & Murant, Ann. appl. Biol. 89: 237, 1978.
- Esau, Am. J. Bot. 44: 245, 1957.
- Esau & Hoefert, Virology 48: 724, 1972.
- Eskandari, Sylvester & Richardson, Phytopathology 69: 45, 1979.
- Falk & Anderson, Phytopathology 77: 1705 (Abstr. no. 141), 1987.
- Falk & Duffus, Phytopathology 74: 1224, 1984.
- Falk, Duffus & Morris, Proc. Am. Phytopath. Soc. 5: 148, 1977.
- Falk, Duffus & Morris, Phytopathology 69: 612, 1979.
- Faoro, Amici & Tornaghi, J. Submicroscop. Cytol. 10: 126, 1978.
- Fargette, Lister & Hood, Pl. Dis. 66: 1041, 1982.
- Francki, Milne & Hatta, Atlas of Plant Viruses, Vol. I, p. 137, CRC Press, Boca Raton, 222 pp., 1985.
- Frazier, Phytopathology 41: 221, 1951.
- Freitag & Severin, Hilgardia 16: 375, 1945.
- Gerlach, Miller & Waterhouse, Barley Yellow Dwarf Newsl. 1: 17, 1987.
- Gibson, in Aphids as Virus Vectors, p. 361, ed. K. Harris & K. Maramorosch, Academic Press, New York, 559 pp., 1977.
- Gibson, Rice & Sawicki, Ann. appl. Biol. 100: 49, 1982.
- Gildow, Phytopathology 72: 1289, 1982.
- Gildow, Phytopathology 75: 292, 1985.
- Gildow, Current Topics in Vector Research 4: 93, 1987.
- Gildow & Rochow, Virology 104: 97, 1980a.
- Gildow & Rochow, Phytopathology 70: 1013, 1980b.
- Gildow & Rochow, Pl. Dis. 67: 140, 1983.
- Gildow, Ballinger & Rochow, Phytopathology 73: 1570, 1983.
- Gill, Phytopathology 60: 1747, 1970.
- Gill & Chong, Virology 95: 59, 1979.
- Gill & Chong, Virology 114: 405, 1981.
- Goodman, Watson & Hill, Ann. appl. Biol. 56: 65, 1965.
- Govier, Ann. appl. Biol. 107: 439, 1985.
- Hammond, Lister & Foster, J. gen. Virol. 64: 667, 1983.
- Hampton, Pl. Dis. 67: 1306, 1983.
- Harrison, CMI/AAB Descr. Pl. Viruses 291, 6 pp., 1984.
- Hewings & D'Arcy, Phytopathology 73: 789, 1983.
- Hu, Rochow & Dietert, Phytopathology 75: 914, 1985.
- Hull & Adams, Ann. appl. Biol. 62: 139, 1968.
- Imaizumi & Kubo, Ann. phytopath. Soc. Japan 46: 54, 1980.
- Inouye, Inouye & Mitsuhata, Ann. phytopath. Soc. Japan 45: 70, 1979.
- Iwaki, Roechan, Hibino, Tochihara & Tantera, Pl. Dis. 64: 1027, 1980.
- Jayasena, Hatta, Francki & Randles, J. gen. Virol. 57: 205, 1981.
- Jedlinski & Brown, Virology 26: 613, 1965.
- Jedlinski, Rochow & Brown, Phytopathology 67: 1408, 1977.
- Jensen, Virology 38: 83, 1969.
- Jensen, Phytopathology 62: 587, 1972.
- Johnstone, in The Winston Churchill Memorial Trust - Fellowship Report, 110 pp., 1983.
- Johnstone, Australas. Pl. Path. 13: 55, 1984.
- Johnstone & Duffus, Aust. J. agric. Res. 35: 821, 1984.
- Johnstone, Duffus, Munro & Ashby, Aust. J. agric. Res. 33: 697, 1982.
- Kojima & Yanase, Bull. Fac. Agric. Niigata Univ. 39: 11, 1984.
- Kojima, Shikata, Sugawara & Murayama, Virology 39: 162, 1969.
- Kubo, CMI/AAB Descr. Pl. Viruses 234, 4 pp., 1981.
- Kubo & Takanami, Ann. phytopath. Soc. Japan 44: 398, 1978.
- Kyriakou, Close & Ashby, N.Z. Jl agric. Res. 26: 271, 1983.
- Leonard & Holbrook, Ann. entomol. Soc. Am. 71: 493, 1978.
- MacKinnon, Can. J. Bot. 43: 509, 1965.
- MacKinnon & Lawson, Can. J. Bot. 44: 1219, 1966.
- Martin & Converse, Phytopath. Z. 114: 21, 1985.
- Mayo & Barker, Rep. Scott. Crop Res. Inst., 1983: 186, 1984.
- Mayo, Barker, Robinson, Tamada & Harrison, J. gen. Virol. 59: 163, 1982.
- Milbrath & Duffus, Phytopath. News 12: 170, 1978.
- Miller, Hercus, Waterhouse & Gerlach, Abstr. 7th int. Congr. Virol., Edmonton, Canada, 1987: 299 (abstr no. OP19.2), 1987.
- Miller, Waterhouse & Gerlach, Nucleic Acids Res. 16: 6097, 1988a.
- Miller, Waterhouse & Gerlach, Virology 165: 306, 1988b.
- Montllor & Gildow, Entomologia exp. appl. 42: 63, 1986.
- Mullin, Smith, Frazier, Schlegel & McCall, Phytopathology 64: 1425, 1974.
- Murant & Roberts, Ann. appl. Biol. 92: 343, 1979.
- Namba, Yamashita, Doi & Yora, Ann. Phytopath. Soc. Japan 45: 70, 1979.
- Orlob & Arny, Phytopathology 51: 768, 1961.
- Paliwal, Phytopath. Z. 92: 240, 1978.
- Prentice, Nature, Lond. 158: 24, 1946.
- Rajeshwari & Murant, Ann. appl. Biol. 112: 403, 1988.
- Reddy, Murant, Duncan, Ansa, Demski & Kuhn, Ann. appl. Biol. 107: 57, 1985.
- Roberts, Tamada & Harrison, J. gen. Virol. 47: 209, 1980.
- Rochow, Phytopathology 50: 881, 1960.
- Rochow, Phytopathology 53: 355, 1963.
- Rochow, Phytopathology 59: 1580, 1969.
- Rochow, A. Rev. Phytopath. 10: 101, 1970.
- Rochow, Phytopathology 65: 99, 1975.
- Rochow, Phytopathology 72: 302, 1982.
- Rochow & Carmichael, Virology 95: 415, 1979.
- Rochow & Duffus, Phytopathology 68: 51, 1978.
- Rochow & Duffus, in Handbook of Plant Virus Infections and Comparative Diagnosis, p. 147, ed. E. Kurstak, Elsevier/North-Holland, Amsterdam, 943 pp., 1981.
- Rochow & Muller, Pl. Dis. Reptr 58: 472, 1974.
- Rowhani & Stace-Smith, Virology 98: 45, 1979.
- Sarkar, Virology 70: 265, 1976.
- Scalla & Rochow, Virology 78: 576, 1977.
- Schalter, Qualset & Rutger, Crop Sci. 4: 544, 1963.
- Scheller & Schukle, Entomologia exp. appl. 40: 189, 1986.
- Shepard & Claflin, A. Rev. Phytopath. 13: 271, 1975.
- Shepardson, Esau & McCrum, Virology 105: 379, 1980.
- Shepherd, Francki, Hirth, Hollings, Inouye, McLeod, Purcifull, Sinha, Tremaine & Valenta, Intervirology 6: 181, 1976.
- Smith, Nature, Lond. 155: 174, 1945.
- Smith, Parasitology 37: 131, 1946.
- Spiegel, Cohen & Converse, Acta Hort. 186: 95, 1986.
- Stubbs, Aust. J. biol. Sci. 63: 326, 1948.
- Stubbs, Smith & O'Loughlin, Abstr. 4th int. Congr. Pl. Path., Melbourne, Australia, 1983: 123 (abstr. 488), 1983.
- Sylvester, Phytopathology 38: 429, 1948.
- Sylvester, Phytopathology 44: 219, 1954.
- Sylvester, A. Rev. Entomol. 25: 257, 1980.
- Takanami & Kubo, J. gen. Virol. 44: 153, 1979a.
- Takanami & Kubo, J. gen. Virol. 44: 853, 1979b.
- Thomas, Ann. appl. Biol. 104: 79, 1984.
- Thottappilly, Kao, Hooper & Bath, Phytopathology 67: 1451, 1977.
- Upreti & Nagaich, Phytopath. Z. 71: 163, 1971.
- Van der Broek & Gill, Phytopathology 70: 644, 1980.
- Waterhouse & Gerlach, Abstr. 6th int. Congr. Virol., Sendai, Japan, 1984: 18 (abstr. P32-15), 1984.
- Waterhouse & Murant, Ann. appl. Biol. 97: 191, 1981.
- Waterhouse & Murant, CMI/AAB Descr. Pl. Viruses 249, 4 pp., 1982.
- Waterhouse & Murant, Ann. appl. Biol. 103: 455, 1983.
- Waterhouse, Gerlach & Miller, J. gen. Virol. 67: 1273, 1986.
- Watson, Rep. Rothamsted exp. Stn, 1963: 104, 1964.
- Watson, Serjeant & Lennon, Ann. appl. Biol. 54: 153, 1964.
- Weidemann, Z. angew. Ent. 94: 321, 1982.
- Yu, Pei & Hsu, Acta phytopath. sin. 3: 1, 1957.
- Zhou & Rochow, Phytopathology 74: 1450, 1984.